
{kind=link}
{kind=link}
封育年限对云雾山典型草原植物繁殖与植被更新的影响
[赵凌平1, 2  , 谭世图
, 谭世图1, 2 , 白欣1 , 王占彬1, 2, * ]
, 谭世图]
|
|


作者简介:赵凌平(1983-),女,河南漯河人,讲师,博士。E-mail:zlp19830629@163.com
研究封育年限对黄土高原典型草原繁殖更新的影响,为退化黄土高原植被恢复和生物多样性维持提供一定的理论依据。在云雾山自然保护区的实验区选取不同封育年限区(5,15,23,32 年)和放牧地(对照)草地作为试验样地,采用挖掘取样法进行草地繁殖更新的研究。结果表明:封育年限对后代繁殖更新苗的物种丰富度影响显著( P<0.05)。封育5年草地和封育15年草地的后代更新苗的物种丰富度与放牧地差异不明显( P>0.05),而封育23年草地和封育32年草地的后代更新苗的物种丰富度显著小于放牧地。封育年限对草地后代繁殖更新总密度、无性繁殖密度、根茎型、分蘖型、匍匐型和分枝型的繁殖密度均影响显著( P<0.05)。随着封育年限的增加,繁殖更新总密度和无性繁殖密度均呈递减趋势,根茎型和分蘖型密度均呈先升高后降低趋势,根蘖型和分枝型密度呈先降低后升高趋势,匍匐型的密度呈逐渐增加趋势,无性繁殖密度与有性繁殖密度之比呈先增加后降低趋势。植物群落密度与无性繁殖密度、繁殖更新总密度之间均呈正显著相关。在云雾山典型草原,短期封育有利于植物后代繁殖更新,长期封育反而不利于草地的繁殖更新,对退化草地封育5年最有利于植物的繁殖更新。植物无性繁殖密度的变化直接影响着繁殖总密度的变化,从而影响了植物群落的密度变化。
To provide a theoretical basis for vegetation restoration and the maintenance of biodiversity, we studied the effects of the duration of enclosure on plant propagation and vegetation regeneration in the semiarid perennial steppe of Yunwu Mountain. We compared non-enclosed (grazed) grasslands and grasslands enclosed for 5, 15, 23, 32 years using the field root-digging method to investigate plant reproductive strategies. The results showed that the duration of enclosure significantly affected species richness ( P<0.05). Species richness was significantly higher in grazed sites than in grasslands with 23 and 32 years of grazing exclusion. However, there was no significant difference in species richness among grazed grassland, grassland with 5 years grazing exclusion, and grassland with 15 years grazing exclusion ( P>0.05). The duration of enclosure significantly affected the densities of total offspring, asexual offspring, rhizomes, tillers, stolons, and branches ( P<0.05), however, the densities of sexual offspring and root suckers did not differ significantly among the treatments ( P>0.05). With prolonged duration of enclosure, the total density and asexual reproduction density decreased; the offspring density from rhizomes and tillers first increased and then decreased; the offspring density from root suckers and branches first decreased and then increased; the offspring density from stolons increased gradually; and the ratio of asexual and sexual reproduction densities first increased and then decreased. There were significant positive correlations between vegetation density and asexual offspring density, and between vegetation density and total offspring density. Our results indicate that short-term enclosure was conducive to offspring recruitment, and long-term enclosure was disadvantageous for grassland regeneration. Fencing for 5 years was the most conducive to offspring recruitment and grassland regeneration in the semiarid steppe of Yunwu Mountain. Compared with sexual offspring, asexual offspring contributed more to total offspring density, which directly affects the density of the plant community. The trends in the change in density in the plant community could be predicted from offspring density during the succession process.
暖温性典型草原是以多年生旱生草本植物为主要组成的草地类型, 也是我国黄土高原面积最大的一类。近些年由于全球气候变暖、过度放牧、落后的畜牧业生产经营方式导致草地退化严重, 草场产量和质量下降, 水土保持功能减弱, 草地生态环境恶化相当严重[1, 2]。为了恢复退化草地生态系统, 维持物种多样性, 对退化草地实行围栏封育已成为区域生态恢复与重建的一项主要措施[3]。草地封育是利用草地植物的自我更新能力来恢复退化草原植被的方法, 其具有投资少、见效快的特点, 从而被广泛的采用[4]。封育对黄土高原退化草地的生态保护与恢复起着重要的积极作用, 现已成为该地区最主要的草地管理措施[5]。但根据草地可持续发展原理, 封育时间不应是无限制的[6]。在长期封育不放牧条件下, 枯落物将会在草地上堆积, 达到一定厚度时容易阻碍种子进入土壤, 还易于发生火灾, 一旦发生火灾必将影响草地的物种组成、地表覆盖状况和植被演替进程。因此, 在退化草地上进行禁牧是必要的, 对退化草地实现植被恢复演替具有重要意义, 但是封育多长时间对退化草地是合适的是以后研究的重点。
退化草地围封后, 不仅包含生物因素诸如植物繁殖策略和可塑性的改变, 还包含非生物因素诸如光照、水分和养分特征的变化[7], 二者共同影响着群落的发展动态。退化草地的成功恢复不仅依赖于土壤养分的供给能力, 还取决于植被繁殖更新能力。可利用繁殖体大小和繁殖更新能力大小为退化草地的自然恢复提供了内在条件和潜在动力[8]。但许多研究报道只侧重于封育对植被特征、物种多样性和土壤理化性质的影响[9, 10, 11, 12], 而忽略了对植物自然更新方式的作用。繁殖更新是植物生活史中关键的环节之一, 保证了种群的延续, 维持着群落的组成与结构稳定, 对生物多样性的保护也起着重要作用[13, 14]。草地植物自然更新可通过无性繁殖和有性繁殖来实现。有性繁殖可以使物种维持基因的多样性并提高对外部环境的适应性[15], 而无性繁殖能够使物种利用有限的空间与资源环境降低干扰风险[16]。有性繁殖与无性繁殖之间存在权衡关系, 对有限资源相互竞争[17]。不同物种甚至同一物种在不同的干扰方式或生境下繁殖更新方式会改变, 至于草地植物采取何种方式更新主要取决于物种的遗传特性和对外界环境压力的适应性[18]。在封育自然恢复过程中, 有性繁殖和无性繁殖在演替过程中发挥的作用有多大?在恢复演替序列上植物的繁殖分配又是如何变化的?这些科学问题的回答将有助于典型草原植物生态恢复理论的纵深发展。因此封育年限对草地动态变化的影响和机制研究, 仅从地上植被和土壤营养的角度研究可能不够全面, 需要从繁殖策略的角度进行补充研究, 才可能揭示出封育对植被群落动态变化和演替进程的影响机制。为此, 开展了不同封育年限对黄土高原典型草原繁殖更新的影响研究, 旨在为退化黄土高原植被恢复、群落演替动态预测和生物多样性维持提供一定的理论依据。
研究区位于宁夏回族自治区固原市东北约45 km的云雾山国家自然保护区(36° 13'-36° 19' N, 106° 24'-106° 28' E)。该地区为典型干旱半干旱黄土丘陵低山区, 属暖温带大陆性季风气候, 夏季炎热短暂, 冬季寒冷漫长。海拔在1800~2100 m, 年平均温度6.7 ℃, 年平均蒸发量1400 mm左右, 年平均降雨量440.5 mm, 少而集中且有极大的年际变化, 每年7月到9月降水量占全年的65%~85%。研究区≥ 10℃的年积温为2259.7 ℃, 无霜期为112~140 d。研究区的地带性土壤为黄土, 泥沙含量64%~73%, 粘土含量17%~20%。土壤中的水分主要来自大气降水, 土壤补水能力差, 土质疏松, 抗侵蚀能力弱。草地是半干旱草原, 植被以针茅属(Stipa)为主。
繁殖更新调查于2014年8月在云雾山草原自然保护区实验区内进行。选取封育5, 15, 23, 32年草地和放牧地 (对照即封育0年)作为试验样地。放牧地为自然放牧状态, 放牧强度为中度放牧(4.6羊单位/hm2)。在每个样地随机选择5个样区, 间距100 m左右, 每个样区随机设置5个25 cm× 25 cm 的样方, 样方间隔至少5 m。采用单位面积挖掘取样法, 同时参照Welling等[19]的方法进行有性繁殖和无性繁殖的调查。有性繁殖是以实生苗的出现来确定, 无性繁殖是以分株或分蘖苗等营养枝的出现来确定。根据幼苗地上部分、根的形态特征和所附着的种壳来确定实生苗的物种。根据新营养枝形成的特点可迅速确定牧草的无性繁殖。在该区, 营养繁殖的器官主要有匍匐茎、分蘖节和根茎等, 根据其将无性繁殖进一步分为5类:根蘖型、匍匐型、根茎型、分枝型和分蘖型, 偶尔出现的其他类型在本研究中忽略不计。在每个样方内, 记录所有植株个体(包括成年植株和幼苗)并统计个数, 然后根据实生苗和萌蘖苗进行分类, 记录所有有性繁殖和无性繁殖的数量。
地上植被调查于2014年7月进行。在每个样区随机设置5个50 cm× 50 cm 的样方进行地上植被调查, 样方间隔至少5 m。详细记录每个样方中的植被总盖度、种类组成、每种植物的个数。
用变异系数(coefficient of variation)来衡量后代繁殖更新物种丰富度的空间异质性。
变异系数(C· V)的计算公式为: C· V=
式中:S为标准差;
用单因素方差分析(ANOVA)比较分析了不同封育年限对后代繁殖更新物种丰富度、有性和无性繁殖密度及权衡关系的影响。用Pearson 相关分析分析繁殖更新密度与地上植被密度的关系。P< 0.05表示差异显著, 以上所有分析均在SPSS 16.0软件里进行。
随着封育年限的延长, 地上植被群落演替的方向为:扁穗冰草+火绒草-杂草类群落→ 扁穗冰草+本氏针茅-杂类草群落→ 本氏针茅+白莲蒿-百里香群落→ 本氏针茅+赖草-火绒草群落→ 本氏针茅群落。本研究共记录到44种植物, 其中多年生草本植物为39种, 一年生草本为4种, 灌木仅出现1种。物种隶属于18个科, 菊科和禾本科植物最多, 分别为10和10种。在放牧地, 更新苗出现37种, 随着群落封育演替进程, 更新苗分别出现38, 30, 25和24种。在封育过程中, 消除了家畜的践踏和采食, 牧草形成了大量的种子, 但实际调查中发现形成实生苗的物种较少, 主要以耐阴低矮的杂类草为主。在该区, 多年生禾本科植物的无性繁殖方式以分蘖为主, 个别植物还以根茎方式繁殖, 例如扁穗冰草和散穗早熟禾。菊科植物的无性繁殖方式以分枝为主, 个别植物还以根蘖、匍匐等方式进行扩展。鳞茎等其他繁殖方式在黄土高原典型草原较少出现(表1)。
| 表1 典型草原物种的繁殖方式 Table 1 Plant reproductive modes in typical steppe |
由表2可知, 在放牧地, 幼苗的物种丰富度变化范围是5~15, 变异系数最小, 为29.17%。在封育15年草地, 幼苗的物种丰富度变化范围是5~12, 变异系数最大, 为37.62%。可见, 对退化草地进行封育后, 明显提高了后代更新苗的空间异质性。封育年限对后代繁殖更新苗的物种丰富度影响显著(P< 0.05)。封育5年草地和封育15年草地的后代更新苗的物种丰富度与放牧地差异不明显(P> 0.05), 而封育23年草地和封育32年草地的后代更新苗的物种丰富度显著小于放牧地, 说明短期放牧未显著改变后代更新苗的物种多样性, 而长期封育则显著降低后代更新苗的物种多样性。长期封育措施减少了一些物种繁殖更新的机会, 而适度放牧能使更多物种有机会进行繁殖更新。长期封育使群落中的优势种长期占据光照、水分、空间资源, 而使其他物种进行繁殖更新的机会较少, 只有一些耐阴的伴生种和偶见种出现。放牧措施抑制了高大草层的发育, 促进了下繁禾草、低矮豆科牧草和杂类草的发育, 并渐渐取得优势, 因此更多物种有机会繁殖更新。
| 表2 不同封育年限草地后代繁殖更新物种丰富度的变化 Table 2 Offspring diversity in grasslands with different enclosure durations |
封育措施对草地植物后代繁殖更新总密度和无性繁殖密度均有显著影响(P< 0.05), 而对有性繁殖密度影响不显著(P> 0.05)。随着封育年限的增加, 繁殖更新总密度呈递减趋势。与放牧地相比, 封育5年草地的繁殖更新总密度显著增加。但到了封育15年和23年, 繁殖更新总密度与放牧地差异不显著。封育32年草地的繁殖更新总密度显著低于其他样地。随着封育年限的增加, 无性繁殖密度的变化趋势与繁殖更新总密度的变化趋势一致。封育5年草地的无性繁殖密度显著高于其他样地, 封育32年草地的无性繁殖密度显著低于其他样地, 而封育15年草地和封育32年草地的无性繁殖密度与放牧地差异不显著。可见, 封育一定时期有利于植被更新, 长期封育草地不仅不利于植被更新, 反而对其起到阻碍作用。长期封育主要是通过降低无性繁殖密度来降低草地的后代繁殖更新密度。在云雾山典型草原, 主要是以多年生牧草占优势, 草地植被的更新主要依靠无性繁殖, 封育措施对草地植被更新的影响也主要是调节无性繁殖特征来进行的(表3)。
| 表3 不同封育年限草地有性繁殖和无性繁殖密度的变化 Table 3 Sexual and asexual offspring density in grasslands with different enclosure durations (mean± SE) No./m2 |
根据营养繁殖器官, 将牧草的无性繁殖进一步划分为5类, 用单因素方差分析不同封育年限对各种类型密度的影响, 结果表明封育年限对根茎型、分蘖型、匍匐型和分枝型的繁殖密度影响显著(P< 0.05), 而对根蘖型的繁殖密度影响不显著(P> 0.05)。随着封育年限的延长, 根茎型和分蘖型密度均呈先升高后降低趋势, 根蘖型和分枝型密度呈先降低后升高趋势, 而匍匐型的密度则呈逐渐增加趋势。与放牧地相比, 根茎型和分蘖型的密度在草地封育5年后显著增加, 封育15年后降低到与放牧地差异不显著。根茎型密度在封育23年和封育32年后显著降低, 而分蘖型密度在封育23年后虽然也呈下降趋势, 但下降不显著, 封育32年则显著低于封育23年草地。与放牧地相比, 封育5年草地的匍匐型密度未显著增加, 但到了封育15年显著增加。放牧地的分枝型密度显著高于其他样地, 封育32年草地的分枝型密度显著高于封育5年、封育15年和封育23年草地, 而封育5年、封育15年和封育23年草地之间的分枝型密度差异不显著(表4)。可见, 在典型草原, 合理放牧条件下牧草的根茎繁殖能力、分蘖能力和分枝能力较强。短期封育后牧草的根茎繁殖能力和分蘖能力增强, 而分枝能力减弱。长期封育后, 匍匐型的牧草繁殖更新能力增强。分蘖型牧草的后代繁殖更新在该区一直占优势地位, 根茎型和分枝型次之, 匍匐型和根蘖型较小。
| 表4 不同封育年限草地不同类型无性繁殖密度的变化 Table 4 Density of different offspring types in grasslands with different enclosure durations (mean± SE) No./m2 |
用无性繁殖密度与有性繁殖密度之比来分析封育和放牧对有性繁殖和无性繁殖权衡关系的影响。方差分析结果表明封育年限对无性繁殖密度与有性繁殖密度之比影响显著(P< 0.05), 说明在黄土高原典型草原地区, 封育年限对有性繁殖和无性繁殖权衡关系有明显影响。二者之比随着草地封育年限的延长先升高后降低, 说明短期封育牧草更多会选择无性繁殖, 长期封育后物种有性繁殖方式有所增加(表5)。
| 表5 不同封育年限草地无性繁殖与有性繁殖密度之比的变化 Table 5 Ratio of asexual to sexual density in grasslands with different enclosure durations |
用Pearson 相关分析分析繁殖更新密度与植物群落密度的关系, 结果表明:植物群落密度与有性繁殖密度之间关系不显著(P> 0.05), 而与无性繁殖密度之间呈显著正相关(r=0.367, P< 0.05), 即随着无性繁殖密度的增加, 群落密度增加(图1)。植物群落密度与繁殖更新总密度之间呈显著正相关(r=0.480, P< 0.05), 即随着繁殖总密度的增加, 群落密度增加(图2)。
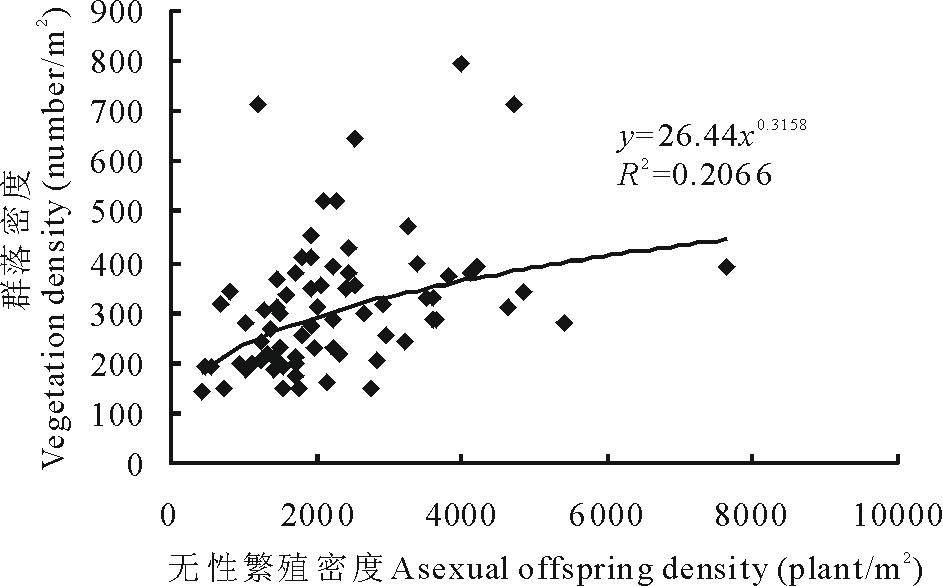 | 图1 无性繁殖密度与植物群落密度的关系Fig.1 Relationship between asexual offspring density and vegetation density |
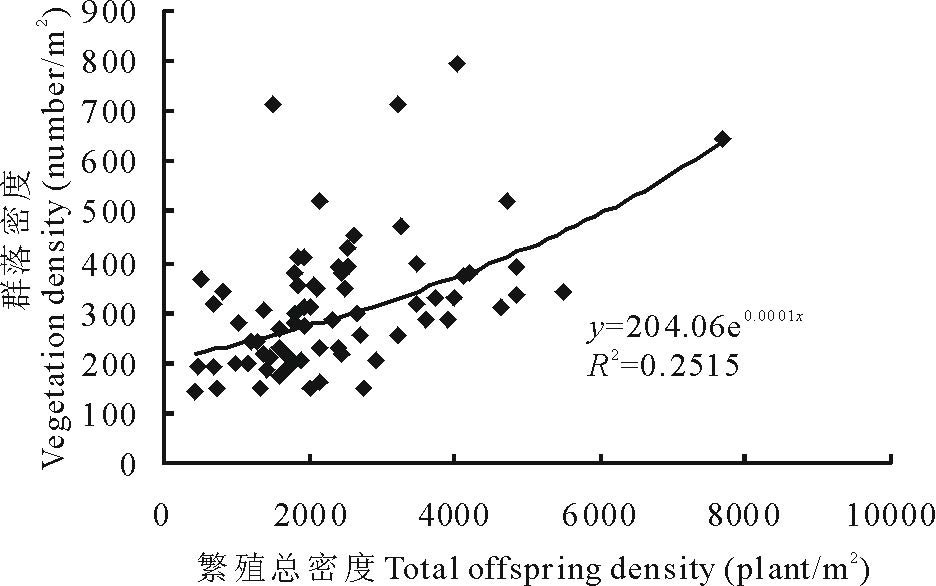 | 图2 繁殖总密度与植物群落密度的关系Fig.2 Relationship between total offspring density and vegetation density |
在黄土高原典型草原群落中, 优势物种是具有根茎或分蘖枝的多年生禾本科植物, 繁殖更新方式主要以无性繁殖为主, 有性繁殖为辅。大多数牧草不是每年都进行种子繁殖, 而常常是每隔一年或数年进行一次, 并根据自然环境条件的适应性而定。在封育草地, 随着封育年限的延长, 以本氏针茅、大针茅等多年生禾本科牧草为主的优势种与其他伴生种、偶见种适应性不断提高, 各项生长指标稳定增长, 然而植物种群数量的变化主要体现在营养繁殖。虽然多年生草本植物每年亦产生大量种子进入到繁殖库中, 但真正萌发长成新植株的数量却很少[20], 这可能因为植物虽然形成大量种子, 但由于种间竞争、生长环境恶劣、春季干旱的环境[21] 、种子形态特征、枯落物厚度、幼苗的竞争力较差[22]等原因, 无性繁殖的分株比有性繁殖的幼苗更易成活。Grime[23] 指出在植被和枯落物茂密繁多、种子繁殖受阻时, 营养繁殖往往容易成功。Armstrong[24]模拟了多年生根茎植物向不同生活史功能过程的生物量分配格局, 发现最佳的资源分配是将1/3的维持后能量分配于种子生产, 而将2/3的维持后能量分配于根茎的生长。因此为了分摊风险, 植物更多地选择无性繁殖来竞争有限资源。但有性繁殖不会被完全放弃, 因为它对种子的扩散具有重要意义。
在黄土高原典型草原, 封育措施对草地繁殖更新的影响较大, 而这种影响在不同封育年限草地中的表现不同。在本试验中由于放牧地采用了适度放牧的措施, 促进了植物的繁殖, 使物种的多样性维持在较高水平, 这符合“ 中度干扰理论” , 即自然干扰下群落优势种竞争力减弱使伴生种和偶见种增加[25]。在适度放牧条件下, 家畜的采食可以刺激更新芽的发育, 也有利于种子进入土壤种子库中补充到幼苗库。在短期封育草地中, 剔除了放牧家畜的啃食与践踏, 退化草地的土壤条件明显改善, 多年生禾草的优势地位还未突显, 分蘖型、分枝型和根茎型牧草的繁殖更新能力明显提高, 因此多年生禾草与多年生杂类草的无性繁殖更新密度显著增加。但长期封育草地不仅没有给牧草的繁殖更新提供有利条件, 反而抑制了牧草的繁殖更新, 特别是抑制了分蘖型和根茎型牧草的繁殖更新, 降低了后代繁殖更新的物种丰富度和密度。这可能是在长期封育环境中, 植物长期处于强烈竞争状态, 多年生草本占主导地位, 劣势种逐渐退出, 枯落物厚度增加, 抑制了植物的再生和幼苗的形成, 不利于草地的繁殖更新。
本研究表明封育年限对克隆植物的有性繁殖与无性繁殖之间的权衡关系影响显著, 放牧和短期封育条件下牧草更多会选择无性繁殖, 长期封育后克隆植物更倾向于将更大比例分配于有性繁殖。在严酷的环境下, 植物的有性繁殖会受到抑制, 无性繁殖往往占优势[26, 27]。有些研究表明一些植物随放牧强度的增大有性繁殖减弱, 无性繁殖增强, 例如冷蒿[28]和矮蒿草[29](Kobresia humills)。在短期封育草地中, 退化草地还未完全恢复, 恶劣的环境限制了植物的有性繁殖。草地经过长期封育后, 幼苗比例增加, 特别是低矮的耐阴的杂类草更多选择种子繁殖来更加成功生存的概率。
植物群落密度与更新苗密度之间的关系在草地生态系统中较少报道, 而在森林生态系统中报道较多。在秦岭山地油松次生林群落中, 林分密度对林下乔木幼苗、幼树数量影响极显著。林下更新的幼苗、幼树密度先随林分密度增加呈增加趋势, 但随着林分密度的继续增加, 其密度逐渐减少[30]。在黄土高原森林区人工油松林中, 林下幼苗的更新状况在低密度林地内最好[31]。而本研究表明植物群落密度与无性繁殖密度和繁殖总密度之间的关系均表现出显著正相关, 说明植物群落的密度显著影响繁殖更新的密度, 但二者之间的关系在草地生态系统和森林生态系统表现不一致。在草地生态系统中, 更新方式以无性为主, 而在森林生态系统中, 更新方式以实生为主[30], 这可能是引起二者关系在不同生态系统表现不一致的主要原因。在本研究中, 植物群落密度与有性繁殖密度之间的相关性不显著。
黄土高原典型草原以多年生牧草占优势, 植被更新主要以无性繁殖为主, 以有性繁殖为辅。其中, 多年生牧草主要依靠分蘖、根茎和分枝等方式来进行无性繁殖。封育年限显著影响草地繁殖更新的物种丰富度、总密度和无性繁殖的密度(P< 0.05)。随着封育年限的增长, 封育草地更新苗的物种丰富度呈先升高后下降趋势, 繁殖更新总密度和无性繁殖密度呈递减趋势。草地群落的繁殖更新在短期封育后达到了最高水平, 长期封育反而不利于后代更新。随着封育年限的延长, 根茎型和分蘖型密度均呈先升高后降低趋势, 根蘖型和分枝型密度呈先降低后升高趋势, 而匍匐型的密度则呈逐渐增加趋势。地上植被密度与无性繁殖密度、繁殖更新总密度之间均呈正显著相关。封育年限对地上植被的影响可通过植物后代的繁殖更新调节, 但影响机制有待进一步研究。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|