






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
植物中钙依赖蛋白激酶(CDPK)的研究进展
[武志刚 , 武舒佳, 王迎春, 郑琳琳
, 武舒佳, 王迎春, 郑琳琳* ]
, 武舒佳, 王迎春, 郑琳琳]
|
|


作者简介:武志刚(1988-),男,内蒙古包头人,在读博士。E-mail:736210941@qq.com
Ca2+是植物细胞信号转导中重要的第二信使,在植株受到外界逆境胁迫时,细胞中的钙信号经钙传感蛋白(CaMs、CaMLs、CBLs和 CDPKs)传递到下游组分引起相关基因的表达,从而响应环境的各种变化。钙依赖性蛋白激酶(CDPK)是Ca2+传感器,可以将细胞内Ca2+信号传递到可以与14-3-3蛋白相互作用的靶蛋白从而刺激靶蛋白发生磷酸化。CDPK在植物生长的诸多方面发挥关键作用,如花粉管的伸长、植物的生长发育以及对生物胁迫、非生物胁迫的应激反应。大多数CDPK具有明显的亚细胞分布,使它们能够“感受”局部Ca2+浓度的变化并与其靶细胞发生特异性作用。14-3-3蛋白是一类通过磷酸化激活的真核蛋白,它可以与不同靶蛋白相互作用,调控植物重要的生理生化过程。近年来研究发现,14-3-3蛋白作为CDPK的靶蛋白和CDPK交叉磷酸化形成CDPK/14-3-3复合物,进一步调节植物初级代谢、开花和激素合成。将从CDPK的结构、亚细胞定位、靶蛋白、生物学功能,特别是CDPK与14-3-3之间的交叉调节及其在植物信号通路中的协同作用方面进行全面的综述,旨在为未来CDPK研究提供参考和新的方向。
Ca2+ is an important secondary messenger in signal transduction in plant cells. When plants are exposed to fluctuating environmental conditions Ca2+ signals are perceived and decoded by Ca2+ sensors (CaMs, CaMLs, CBLs and CDPKs) to elicit the expression level of related genes. Calcium-dependent protein kinase (CDPK) is a Ca2+ sensor playing a pivotal role in plant development, pollen tube elongation and responses to abiotic and biotic stimuli. It has the unique ability to directly transmit cytolic Ca2+ signals to downstream phosphorylation events in diverse substrates which can mediate interaction with 14-3-3 proteins to modulate protein functions. Most CDPKs have significant subcellular distribution, allowing them to “feel” local Ca2+ concentration and to act specifically with their target cells. 14-3-3 proteins are highly conserved in eukaryotic cells, which in most cases need to bind to different targets and be phosphorylated to modulate their activity. Through protein-protein interactions, 14-3-3 proteins are involved in many significant physiological processes in plants. Recent studies have revealed that the role of CDPK in phosphorylating sites in mediating 14-3-3 protein binding to form CDPK/14-3-3 complex, and have also highlighted the role of the CDPK/14-3-3 complex in regulating primary metabolism, plant hormone synthesis and flowering. In this paper, CDPK structure, subcellular localization, target protein, biological functions, especially the cross regulation between CDPK and 14-3-3 and their synergistic effect in plant signaling pathway will be discussed in depth. Our study aims are to provide a reference and indicate new directions for future CDPK research.
植物中存在着复杂的信号转导网络, 用于调控植物的新陈代谢, 以应对不断变化的生存环境。Ca2+作为重要的第二信使, 经Ca2+受体蛋白、钙调素(CaM)和钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)等协同作用[1], 将钙信号向下游传递并级联放大, 促进响应蛋白的产生, 从而调控植物的生长发育以及免疫应答和胁迫响应。
CDPK是目前植物中研究较多、了解较为清楚的一类丝氨酸/苏氨酸蛋白激酶, 在植物发育及其对生物和非生物胁迫信号转导的应答中执行不同的生物学功能[2, 3, 4]。基因组数据分析显示, 在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、小麦(Triticum aestivum)、玉米(Zea mays)及杨树(Populus tomentosa)中分别含有34、31、20、40、20个CDPK基因[5, 6]。后续研究发现CDPK同样存在于绿藻类、卵菌和纤毛虫、顶覆虫等部分原生动物中。CDPK在不同组织中的表达、亚细胞定位、作用底物以及与不同蛋白体形成复合物等方面的差异, 决定了该家族成员功能的差异, 因此, 本研究拟从这些方面对CDPKs进行全面的概述。
CDPK首次发现于豌豆(Pisum sativum)中, 第1次从大豆(Glycine max)中得到纯化和鉴定。与其他钙结合蛋白不同, CDPK是单肽链结构。如图1所示, CDPK具有4个典型的结构域, 从N端到C端依次为N-末端的可变域、催化域(激酶域)、连接域(自抑制域)和调控域(类钙调素域/CaM-LD)。
 | 图1 CDPK的结构特征Fig.1 Structural features of CDPK |
N-末端可变域序列相似性低, 保守性差[7]。多数 CDPK 在N-端可变域含有与膜定位相关的豆蔻酰化位点(第2位的甘氨酸残基)以及棕榈酰化位点(第4或第5位的半胱氨酸残基)[8], 参与CDPK的膜结合过程。此外, N-末端可变域还在调节底物特异性过程中扮演关键的角色[9, 10]; 催化域也称激酶域, 具有典型的Ser/Thr蛋白激酶的保守序列, 同源性较高[11], 该区域突变会导致催化活性丧失; 连接域高度保守, Liese等[7]的研究表明, 连接域功能上相当于一种假底物, 与催化域结合保证CDPK处于非活跃状态, 因此也叫自抑制域; C-末端调控域上具有类似钙调素的结构, 因此CDPK的激活依赖于钙离子而不依赖于钙调素。调控域包含4个可与钙离子结合的EF-手性结构, 是Ca2+和CDPK的结合位点[12]。根据对Ca2+的亲和性不同将其分为两个球形结构, 分别为N-lobe和C-lobe[13], N-lobe对Ca2+亲和性低, C-lobe对Ca2+亲和性高, 当Ca2+浓度低时与C-lobe结合, 当Ca2+浓度高时与N-lobe结合, 与连接域相互作用, 导致调控域发生构象变化, 解除自抑制。
CDPK在植株中分布极其广泛, 器官水平上, 在植株的根、茎、叶、花、果实及种子中均能检测到基因的表达; 细胞水平上, CDPK基因普遍存在于分生组织细胞、木质部细胞、保卫细胞、花粉母细胞及胚胎细胞中。CDPK的家族成员的表达特点不尽相同, 有些可以在植物的大多数器官或组织中表达, 如烟草(Nicotiana tabacum)NtCPK1在叶片中不表达, 在根、茎和花中均有表达[14]; 拟南芥的AtCPK12在根、茎、叶、花和成熟果荚等组织中均有表达, 但在干种子中不表达[15]; 油菜(Brassica campestris)BnCDPK1 在根、茎、叶、花和种子中都有表达, 而有些却只在特定的组织表达, 如AtCPK16、AtCPK17、AtCPK25、AtCPK34只在花粉管中表达[16]; 此外, CDPK在不同器官中表达丰度也存在差异, 油菜BnCDPK1在叶片中表达量最高, 其次为茎、根和种子, 在花中的表达量最低[17]。
通过亚细胞定位研究表明, CDPK可定位于细胞质膜、内质网膜、胚乳贮藏囊泡、细胞骨架、线粒体、染色质、细胞核、细胞质、过氧化物酶体、油体及液泡膜等部位, N-末端可变域是CDPK亚细胞定位和功能的关键[18]。利用CDPK与绿色荧光蛋白(GFP)融合表达的方法, 发现拟南芥CDPK几乎全部定位于质膜, 少数定位于核。大多拟南芥CDPK具有专一的膜结合位点, 即N-末端可变域的酰基化位点, 其中豆蔻酰化位点与靶细胞膜形成一个不可逆转的松散的结合, 而棕榈酰化位点则具有可逆的稳定的膜锚定结合[19]。豆蔻酰化位点上的甘氨酸突变为丙氨酸(如AtCPK2、AtCPK3、AtCPK5、AtCPK6、AtCPK9和AtCPK13)使CDPK的膜结合能力降低[20], 棕榈酰化位点突变则使亚细胞定位由膜向核转变。另外, CDPK的亚细胞定位也受非生物胁迫的影响, 如冰草(Agropyron cristatum)McCPK1响应低温胁迫而改变亚细胞定位, 由定位于质膜向定位于细胞核、内质网(ER)和肌动蛋白丝转变[21]。
了解CDPK在植物体内作用的靶蛋白, 对研究其功能具有重要意义。到目前为止, 已发现多种CDPK作用的底物(表1), 它们参与多种细胞过程, 如初级和次级代谢、胁迫响应、离子和水分运输、转录过程、信号转导途径等。
| 表1 已有文献报道的CDPK在植物体内作用的靶蛋白及其调控作用 Table 1 The target protein and its regulatory role of CDPK |
在上述CDPK的靶蛋白中, 14-3-3蛋白因其与CDPK之间的交叉磷酸化作用受到学者的广泛关注。14-3-3蛋白既与磷酸化的CDPK结合也可被CDPK磷酸化, 在真核生物中通过磷酸化及与蛋白质互作调节多种生物学过程。
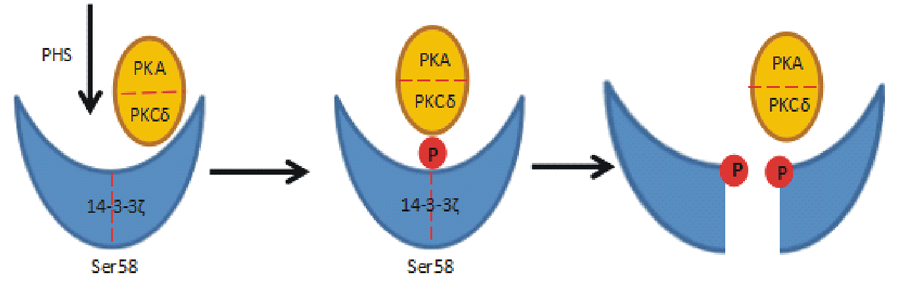
14-3-3蛋白对CDPK的活性有直接调节作用。拟南芥AtCPK1是第1个被发现的受14-3-3蛋白调控的CDPK[45], 但在体外实验中, AtCPK1未能与14-3-3ω 蛋白相结合, 因此推测AtCPK1和14-3-3ω 之间的相互作用可能发生在胞内[46], 作用位点仍有待鉴定。14-3-3蛋白除了参与CDPK活性的直接调节外, 还可以控制CDPK的稳定性。拟南芥AtCPK3已被证实为14-3-3的相互作用蛋白[47, 48], 尽管体外实验显示14-3-3蛋白并不影响AtCPK3的激酶活性, 但近期研究发现, 饥饿诱导胁迫的AtCPK3在与14-3-3蛋白质相互作用丧失后被选择性切割[49], 证明拟南芥细胞内的14-3-3与AtCPK3的互作可以稳定AtCPK3的活性。此外, 在拟南芥细胞程序性死亡过程中, 坏死性真菌产生的毒素FB1(Fumonisin B1)诱导拟南芥中程序性死亡因子PHS(phytosphingosine)表达, PHS水平的增加导致胞内Ca2+升高, 从而激活AtCPK3活性并磷酸化14-3-3蛋白质, 14-3-3的磷酸化使得14-3-3/AtCPK3复合体解离, AtCPK3被切割[50, 51, 52], 这一过程进一步证实了14-3-3蛋白对CDPK具有保护作用。
14-3-3蛋白不仅结合并调节磷酸化后的CDPK, 它们自身也被磷酸化。动植物14-3-3s的各个位点都有可能被磷酸化, 磷酸化在调节14-3-3与靶蛋白的相互作用过程中发挥了极其重要的作用[53, 54]。在动物细胞中, 程序性死亡因子PHS诱导两种不同的蛋白激酶PKA(protein kinase A)和PKCδ (protein kinase Cδ )识别并磷酸化14-3-3ζ 二聚体界面处的丝氨酸残基(Ser58), Ser58磷酸化破坏14-3-3s的二聚体结构, 从而完成了细胞凋亡过程(图2); 在植物细胞中, 与过氧化物酶体结合的AtCPK1和AtCPK3、AtCPK6、AtCPK8、AtCPK24、AtCPK28均可在多个位点磷酸化蛋白14-3-3χ 和14-3-3ε [24], 其中对14-3-3χ 和14-3-3ε 蛋白磷酸化最快速的是AtCPK3和AtCPK28, 因为二者N-末端可变域序列具有高度的同源性。
拟南芥具有13个14-3-3蛋白编码基因, 在植物发育的不同阶段和不同组织中差异表达。CDPK/14-3-3复合体在植物信号通路中协同作用, 共同调控代谢、激素信号传导和气孔运动, 以及对非生物和生物胁迫的响应等重要的生理生化过程[55, 56]。
当光合作用不活跃时, 拟南芥硝酸还原酶NR(nitrate reductase)在丝氨酸残基(Ser534)处被AtCPK17或AtCPK28磷酸化, 14-3-3蛋白通过结合叶片中磷酸化的NR, 阻止亚硝酸盐的产生, 使其不能进一步还原成铵, 最终导致NR失活[24, 42]。菠菜(Spinacia oleracea)硝酸还原酶SoNR则在丝氨酸残基(Ser543)处被相关蛋白激酶SnRK(SNF1-related protein kinase)磷酸化后与抑制性蛋白14-3-3结合, 由于拟南芥AtCPK3与菠菜SnRK具有非常相似的氨基酸序列, 因此拟南芥AtCPK3能够使菠菜NR磷酸化[41]。14-3-3蛋白也可通过结合被AtCPK3磷酸化的拟南芥6-磷酸果糖-2-激酶F2KP(fructose-2, 6-bisphosphatase)参与碳循环过程[25, 46]。
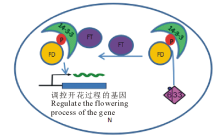
CDPK与赤霉素(GA)信号传导存在紧密的联系。1995年, 在水稻中首次发现由GA诱导的未知的膜定位CDPK[43]。10年后, Abbasi等[56]研究显示, OsCDPK13在转录水平响应GA而被活化; 拟南芥AtCPK28为GA和茉莉酸(JA)的调节剂, AtCPK28突变体改变了维持GA稳态的关键酶的表达, 表现出枝条和叶柄生长缓慢的特性[57, 58]; 烟草中参与GA生物合成的反馈调节因子芽生长抑制因子RSG(repression of shoot growth)被鉴定为NtCDPK1的直接靶标, RSG通过控制GA生物合成基因的转录来维持GA稳态。RSG在细胞质基质和细胞核之间来回移动, 当GA浓度降低时, RSG转入细胞核中激活GA合成基因的转录, 合成的GA诱导细胞质基质中游离Ca2+增加, 从而导致NtCDPK1构象发生改变, 促进RSG在S114片段磷酸化并与14-3-3蛋白结合。NtCDPK1与RSG和14-3-3蛋白质形成异源三聚体并作为支架和媒介促进磷酸化后的RSG与14-3-3形成二聚体, RSG/14-3-3复合体从NtCDPK1脱落后, 细胞质基质中的RSG失活, 核内靶基因的活化被抑制[43, 58, 59](图3)。
 | 图3 CDPK和14-3-3蛋白对GA稳态的调控[52]N: 细胞核Nucleus; C: 细胞质基质Cytoplasmic matrix; P: 磷酸化Phosphorylation.下同The same below.Fig.3 The regulation of CDPK and 14-3-3 protein on GA homeostasis[52] |
在植物乙烯的生物合成途径中, 限速酶1-氨基环丙烷-1-羧酸合酶ACS(1-aminocyclopropane-1-carboxylic acid ACC synthase)家族起到了催化调控的作用。ACS因其C-末端存在不同的序列被分为3种类型, 14-3-3s与所有类型的ACS均可以相互作用[39]。拟南芥AtACS7是一种3型ACS, 可以被AtCPK16在3个位点(Ser216、Thr296和Ser299)处磷酸化, 然后在细胞质基质中与14-3-3ω 直接相互作用形成蛋白复合体[60, 61], AtCPK16和14-3-3ω 的协同作用增加了AtACS7的稳定性。
CDPK是植物所有主要发育过程包括传粉、生长、开花和衰老的调控者。许多拟南芥CDPK(AtCPK2、AtCPK14、AtCPK16、AtCPK17、AtCPK20、AtCPK24、AtCPK26和AtCPK34)主要在花粉中表达, 表明它们参与花粉发育或花粉管伸长。最近的研究表明, 拟南芥中的两种CDPK, 依赖Ca2+的AtCPK11和不依赖于Ca2+的AtCPK24可以形成级联信号, AtCPK11磷酸化AtCPK24 N-末端结构域, 在花粉管特异性钾离子内流通道SPIK(shaker pollen inward K+)定位的质膜处相互作用并抑制其活性, 从而对花粉管伸长进行负调节作用。此外, 其他两种花粉特异性CDPK, AtCPK17和AtCPK34也定位于细胞质膜, 推测可能与花粉伸长过程相关, 研究发现AtCPK17/AtCPK34双突变体在花粉管的顶端极化生长表达上存在缺陷, 而单个的突变则正常表达[64, 65]。
在产生钙信号过程中, 拟南芥内质网膜和细胞质膜上分别具有自我抑制型Ca2+泵ACA2(autoinhibted calcium ATPase 2)和ACA8(autoinhibted calcium ATPase 8), 这两种Ca2+泵调控Ca2+泵入内质网腔或泵出细胞, 维持细胞质基质中较低的Ca2+浓度, 二者均可以直接被CDPK磷酸化。Giacometti等[66]研究表明, AtCPK16定位于质膜, 因此在质膜内磷酸化AtACA8。AtCPK1可以与过氧化物酶体和微粒体结合, 推测与过氧化物酶体相结合的AtCPK1/P复合体可作用于AtACA2和AtACA8; 而ER上可衍生出微粒体, 因此推测与微粒体相结合的AtCPK1/LB复合体可能与AtACA2连接并调控其作用(图5)。
研究表明, CDPK参与植物非生物胁迫反应, 低温[67]、光[68]、干旱[69]、盐害[70]、低渗透[71]、营养饥饿等多种环境因子以及赤霉素GA、生长素、细胞分裂素等激素的诱导都能引起 CDPK基因的特异性表达, 如表2所列。
| 表2 不同植物中响应非生物胁迫的CDPK Table 2 CDPK responsed to abiotic stresses in different plants |
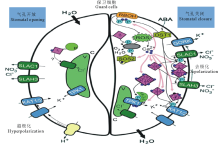
ABA依赖性的气孔闭合是防止干旱胁迫期间水分流失的关键机制[80], 许多CDPK参与到气孔运动的调控过程中[81]。在拟南芥中, 离子通道AtKAT1和AtKAT2以及其他K+转运蛋白具有调节气孔开放的功能[54, 80]。AtCPK1则激活液泡膜上的Cl-通道, 导致Cl-内流; AtCPK1介导的Cl-内流、AtKAT1/AtKAT2介导的K+内流、阴离子通道SLAC1(slowtype anion-associated 1)与SLAH3(SLAC1 homologue 3)介导的阴离子(Cl-、N
AtCPK13是保卫细胞中主要表达的Ca2+敏感性CDPK, 磷酸化AtKAT1和AtKAT2[82], 从而抑制K+内流过程。Sato等[83]的研究表明, AtSnRK2和拟南芥AtCPK4、AtCPK13可以在细胞内磷酸化AtKAT1的C-末端片段T306和T308, 表明拟南芥CDPK和SnRK2之间可能存在一致的靶位点; T306的突变导致AtKAT1和AtKAT2通道活性受损。ABA信号传导产生激活的AtCPK3、AtCPK6、AtCPK21和AtCPK23以及AtOST1的活性氧(ROS)和Ca2+信号, 这些激酶在保卫细胞离子通道的调节中起主要作用, 它们激活AtSLAC1和AtSLAH3阴离子通道, 促进阴离子的流出[84, 85, 86]。此外, AtCPK3、AtCPK4、AtCPK5、AtCPK11和AtCPK29在液泡外磷酸化双孔钾离子通道TPK1(tandem-pore K+ channel 1), 磷酸化的AtTPK1与14-3-3蛋白质形成蛋白复合体介导液泡中的K+流出[87]。质膜质子泵的抑制依赖于Ca2+的阴离子通道的活化导致质膜去极化, 激活保卫细胞外向整流型K+通道GORK(guard cell outward-rectifying K+ channel), K+外流。最终, 保卫细胞内溶质的流出导致其膨胀程度降低, 气孔闭合(图6右)。
到目前为止, CDPK的结构、亚细胞定位以及调控机制等研究已经比较成熟, 越来越多CDPK家族成员的新功能已经被发现, 但其作用底物和抑制剂还有待于进一步的研究, 如弓形虫寄生病的治疗。弓形虫是一种常见的原生动物, 几乎可以感染所有温血动物和人类, 目前尚缺乏有效的治疗药物, 但由于弓形虫中有CDPK, 而人体并无CDPK, 因此CDPK专一性抑制剂可能成为弓形虫治疗的新型药物。此外, CDPK和14-3-3蛋白质之间存在复杂的调控网络。如本研究所述, CDPK不仅磷酸化14-3-3蛋白, 其本身也被14-3-3磷酸化, 尽管近年的多项研究都显示出CDPK/14-3-3复合体在植物活性调控过程中的重要作用, 但CPDKs与14-3-3s通过磷酸化的交叉调节过程仍不清晰, 有待于国内外学者的进一步探索。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|