





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
兰州百合连作障碍效应及机制研究
[黄钰芳1, 2  , 张恩和
, 张恩和1, * , 张新慧3 , 王惠珍2 , 王琦1 , 刘青林1 , 石雨仟1 ]
, 张恩和, 张新慧|
|


作者简介:黄钰芳(1977-),女,甘肃陇南人,讲师,在读博士。E-mail: 1049395950@qq.com
为探讨兰州百合连作障碍效应及作用机制,以正茬及不同连作年限土壤为试验对象,利用盆栽试验,动态研究了连作对百合生长、光合特性及抗氧化酶系统的影响,并利用GC-MS技术分析了正茬及不同连作年限土壤中自毒成分的种类及含量。结果显示,苗期、鳞茎膨大期及成熟期连作对百合的生长均存在抑制作用,且随着连作年限的延长作用程度增强。同一生长阶段,随连作年限的增加,百合植株中CAT和SOD含量逐渐下降,POD活性先升高后降低,MDA含量持续升高。叶绿素总量(Chl)、净光合速率( Pn)、气孔导度( Gs)及蒸腾速率( Tr)呈下降趋势,而胞间CO2浓度( Ci)持续升高。从正茬、连作2年和连作4年土壤中分别鉴定出9种、15种、17种化感自毒物质,主要包括:2,3-丁二醇、间苯二甲酸二辛酯、2,2'-亚甲基双-(4-甲基-6-叔丁基苯酚)和对苯二甲酸二辛酯。其中,对苯二甲酸二辛酯在连作2年和4年土壤中含量均达最高值,分别为33.24%和41.43%。连作通过影响百合植株的光合作用能力,破坏抗氧化酶代谢平衡,从而抑制百合植株的生长,导致连作障碍的发生;连作条件下多种次生代谢物引起的自毒作用是导致兰州百合连作障碍产生的主要原因之一。
, ZHANG En-he, ZHANG Xin-huiTo explore the negative effects of continuous cropping on the growth of Lanzhou lily ( Lilium davidii var. unicolor), we conducted a pot experiment using soil containing fresh Lanzhou lily stubble and soil from fields where Lanzhou lily had been cultivated for 2 or 4 years. The growth indexes, photosynthetic characteristics, and antioxidant enzyme activity of Lanzhou lily plants were evaluated, and the types and contents of autotoxic substances in soil from fields continuously cropped with Lanzhou lily for 2 or 4 years was analyzed by gas chromatography-mass spectrometry (GC-MS). The results showed that the growth of Lanzhou lily was inhibited by continuous cropping at the seedling stage, bulb enlargement stage, and maturity stage, and the inhibitory effect increased with increasing number of continuous cropping years. At the same growth stage, with increased number of continuous cropping years, the activity of catalase and superoxide dismutase decreased, the activity of peroxidase increased and then decreased, the malondialdehyde content increased, the total chlorophyll content, net photosynthetic rate, stomatal conductance, and transpiration rate decreased gradually, and the intercellular CO2 concentration increased. We identified 9 compounds in soil containing stubble, and 15 and 17 compounds in soil where Lanzhou lily had been continuously cropped for 2 and 4 years, respectively. These compounds included 2, 3-butanediol, dioctyl isophthalate, 2, 2'-methylenebis-(4-methyl-6-tert-butylphenol), and dioctyl terephthalate. The highest contents of dioctyl terephthalate were in soil where Lanzhou lily was continuously cropped for 2 and 4 years (33.24% and 41.43%, respectively). Continuous cropping inhibited the growth of Lanzhou lily by affecting its photosynthetic capacity and disrupting the metabolic balance of antioxidant enzymes, ultimately leading to reduced performance under continuous cropping. The autotoxicity of several secondary metabolites that accumulated under continuous cropping was one of the main causes of the reduced performance of Lanzhou lily.
兰州百合(Lilium davidii var. unicolor salisb)是百合科(Liliaceae)百合属(Lilium)川百合的变种, 为多年生鳞茎草本植物, 是我国四大百合品系中唯一可食用的品系, 由于其鳞茎硕大洁白, 质地细腻, 营养丰富, 故被誉为“ 蔬菜人参” [1]。然而, 近年来由于快速发展的兰州百合产业, 造成兰州百合种植区域连作重茬加重, 导致兰州百合产量和品质明显下降, 兰州百合产量由原来的约2.25万kg· hm-2下降到1.50万kg· hm-2, 产品的独头率从原来的70%下降到不足40%[2], 严重影响了兰州百合的市场声誉和农民的经济效益。因此, 连作障碍已成为困扰兰州百合产业持续发展的主要问题。前人对植物连作障碍产生的原因进行了大量研究, 认为植物根系分泌物的累积、土壤微生物种群平衡的破坏及根际微生态平衡的失调等因素是导致植物连作障碍发生的主要原因[3]。然而, 连作障碍发生机理的研究多集中在大豆(Glycine max)、花生(Arachis hypogaea)、黄瓜(Cucumis sativus)、马铃薯(Solanum tuberosum)、地黄(Rehmannia glutinosa)、当归(Angelica sinensis)[4, 5, 6, 7, 8, 9]等作物, 兰州百合障碍效应以及化感自毒物质对于百合植株的生理生态作用及响应机制未见报道。故本试验通过分析不同连作条件下百合植株生长、光合作用能力、抗氧化酶系统及受试土壤中化感自毒物质种类及含量的变化揭示了兰州百合连作障碍效应及其作用机制, 以其为消减和解除兰州百合连作障碍提供一定的科学依据。
试验地位于兰州百合主产地— 兰州市七里河区西果园镇, 海拔1700 m。该地区属兰州市南郊二阴山区, 大陆性半干旱气候, 年平均气温5.8 ℃, 生长期平均232 d, ≥ 10 ℃的年积温为2200 ℃, 无霜期平均135 d, 年均降水量460 mm, 适合兰州百合喜温凉、喜光照、好湿润的生理特征。
于2016年3月底百合春季采收期, 在试验地选择同一海拔, 坡度相近的阳坡收集正茬及不同连作年限土壤, 以当地同年收获的兰州百合种球为受试对象, 进行盆栽试验。
根据土壤来源的不同共设正茬(歇地3年土壤)、连作2年和连作4年3个处理, 分别标记为CK、T1和T2。3种处理土壤速效N分别为:571.0、613.8和140.8 mg· kg-1。速效P分别为:229.6, 23.7和30.4 mg· kg-1。速效K分别为:378.1、324.4和348.5 mg· kg-1, 以最高测定值为依据, 将每一处理中土壤肥力补充至同一水平。试验采用口径31 cm, 底径19 cm, 高25 cm的聚乙烯塑料盆, 每盆装入土壤5 kg。选择包合紧密、基盘健康、大小均一的百合种球, 称重后消毒(在浓度为3 mg· mL-1的高锰酸钾溶液中灭菌10 min, 再用无菌蒸馏水冲洗2~3次), 每盆栽培8株, 待正常生长后定植4株。每处理重复5次, 每重复设计5盆。盆栽随机排列置于室外, 全生育期正常管理。
于苗期(75 d)、鳞茎膨大期(125 d)及成熟期(170 d)分别取样进行百合植株生长指标、光合色素含量及抗氧化酶活性的测定; 于鳞茎膨大期进行光合生理指标测定并分析不同处理中化感自毒物质的种类及含量。
1.3.1 形态指标的测定 采用匀盆的方式, 随机选取各处理盆栽, 用直尺测定其株高与主根长, 用万分之一天平测定根、鳞茎、茎叶鲜重(每处理重复3次, 每重复4株)。
1.3.2 光合生理指标的测定 采用Li-6400便携式光合测定仪(Li-Cor Inc., USA), 人工控制CO2浓度为400 μ mol· mol-1、温度25 ℃、光照强度1200 μ mol· m-2· s-1, 测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci), 每处理3次重复, 每重复记录10个数据。
叶绿素含量测定:分别取各处理受体同一部位成熟叶片, 采用丙酮-乙醇混合液提取法[10]提取色素, 用722PC型分光光度计测定叶绿素a和叶绿素b含量。
1.3.3 抗氧化酶活性的测定 过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)和丙二醛(MDA)含量分别采用硫代硫酸钠滴定法、愈创木酚法、氮蓝四唑(NBT)光还原法和硫代巴比妥酸(TBA)法测定[11]。
1.3.4 化感自毒物质的检测 分别称取各处理新鲜根际土壤500 g, 加适量蒸馏水浸泡24 h, 期间多次震荡。滤过, 滤液以4000 r· min-1离心10 min, 取上清液, 减压抽滤, 滤液加入等体积氯仿萃取5次, 用旋转蒸发仪在4 ℃条件下减压回收氯仿至0.5 mL, 用于GC-MS分析。
GC-MS条件:HP-5MS(30 m× 250 μ m× 0.25 μ m)柱。进样口温度250 ℃; 柱温40 ℃。以5 ℃· min-1程序升温至50 ℃, 保持2 min, 再以5 ℃· min-1程序升温至130 ℃, 保持5 min, 再以5 ℃· min-1程序升温至160 ℃, 保持5 min, 再以5 ℃· min-1程序升温至250 ℃, 保持5 min, 再以5 ℃· min-1程序升温至280 ℃, 保持10 min, 最后以10 ℃· min-1程序升温至300 ℃。载气:He, 流量:1 mL· min-1, 进样量为1.0 μ L。电子轰击源, 轰击电压70 eV, 扫描范围M/Z 30-600 AMU, 扫描速度0.2 S扫全程, 离子源温度230 ℃。
应用NIST11.L质谱数据库计算机检索系统分析质谱图, 进行未知物的鉴定。
利用Excel 2007软件处理原始数据, 通过SPSS 17.0软件, 对数据进行单因子方差分析, 用新复极差法(Duncan)做显著性检验。
由表1可知, 连作可以抑制百合的生长发育, 苗期、鳞茎膨大期和成熟期3个生长阶段, 随着连作年限的增加, 百合植株各生长指标均有所下降。苗期, T1中株高、茎叶重、根长、根重、鳞茎增长量(鳞茎重-种球重)较CK分别下降了7.86%、13.32%、13.13%、23.49%和6.08%, T2中分别下降了12.47%、51.04%、20.20%、37.72%和51.33%, 且T2较CK除根长外, 其余指标均达显著差异(P< 0.05)。鳞茎膨大期及成熟期, 随连作年限的延长, 百合植株各生长指标均持续下降, 并且3处理间差异均达显著水平(P< 0.05)。
2.2.1 连作对百合叶绿素含量的影响 叶绿素含量的高低是衡量叶片对光能吸收能力的指标之一[12]。由图1(A)可知, 3个生长期, T1较相应CK叶绿素总量分别下降了14.64%、20.82%和30.16%, 其中鳞茎膨大期时差异显著(P< 0.05); T2较相应CK分别下降了28.21%、38.92%和60.32%, 差异均显著(P< 0.05), 且鳞茎膨大期时与T1间差异也呈显著水平(P< 0.05), 可见连作对百合植株叶绿素总量有抑制作用, 随着连作年限的增加抑制作用加强, 并且这种作用伴随在整个生育期内。
Chla/Chlb值是反映类囊体膜垛叠程度的敏感指标, Chla/Chlb值的变化能够反映叶绿体捕光能力的高低及能量传递速率的快慢[13]。由图1(B)可知, 随着生育期的发展, 连作处理中Chla/Chlb值呈持续上升趋势; 而同一生育阶段, 随着连作年限的增加Chla/Chlb值同样呈升高趋势, 表明与正茬相比, 连作能够降低百合类囊体膜的垛叠程度, 从而降低叶绿体的捕光能力及能量传递速率。
| 表 1 连作对百合植株生长指标的影响 Table 1 Effect of continuous cropping on growth indexes of lily plant |
 | 图1 连作对百合不同生育时期叶绿素总量(A)及Chla/Chlb(B)的影响 不同小写字母表示差异显著(P< 0.05), 下同。Fig.1 Effect of continuous cropping on the sum chlorophyll and Chla/Chlb of lily at different growth stages The different lowercase letters mean the significant differences at P< 0.05, the same below. |
2.2.2 连作对百合光合作用的影响 光合作用是绿色植物生物产量和经济产量形成的源泉, 光合能力则是反映植株与周围环境适应程度的重要生理指标[14]。由表2可知, 鳞茎膨大期, 与CK相比, T1中净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)分别下降28.03%、14.25%和23.56%, T2中分别下降40.59%、23.97%和40.84%, 而两处理中胞间CO2浓度则分别增加了18.00%和21.20%, 3处理间各指标差异均达显著水平(P< 0.05)。可见, 连作可以降低百合叶片的光合作用, 并且随着连作年限的增加, 光合能力逐渐下降。
| 表 2 连作对百合光合作用的影响 Table 2 Effect of continuous cropping on photosynthesis of lily |
由图2~图4可知, 不同连作年限下百合叶片POD活性随着生育阶段的变化均表现为苗期> 鳞茎膨大期> 成熟期, CAT和SOD活性均呈现出鳞茎膨大期最大, 苗期次之, 成熟期最小。相同生育阶段, 随着连作年限的增加, 百合叶片CAT和SOD活性均呈现持续下降趋势, POD活性却表现为先升高而后降低。鳞茎膨大期, T1中POD活性较对照增加了6.68%, T2降低了4.28%, 而CAT和SOD活性, 在两处理中均降低, 其中T1较对照分别下降了31.61%和49.04%, T2中分别下降了38.75%和59.99%, 差异均显著(P< 0.05)。由此看出, 连作可以影响百合叶片中抗氧化酶的活性, 并且对于不同保护酶活性其影响趋势和作用程度存在一定的差异性。
 | 图2 连作对百合不同生育时期叶片CAT活性的影响Fig.2 Effect of continuous cropping on CAT activity in leaves of lily at different growth stages |
 | 图3 连作对百合不同生育时期叶片POD活性的影响Fig.3 Effect of continuous cropping on POD activity in leaves of lily at different growth stages |
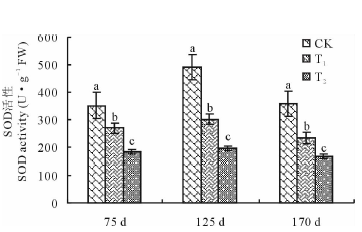 | 图4 连作对百合不同生育时期叶片SOD活性的影响Fig.4 Effect of continuous cropping on SOD activity in leaves of lily at different growth stages |
由图5可知, 3个生长阶段, T1、T2处理中MDA含量与相应对照相比分别增加了30.28%和62.78%, 49.89%和101.94%, 31.98%和64.34%, 差异显著(P< 0.05)。可以看出, 随着连作年限的延长, 生育进程的推进, 百合叶片中MDA含量均呈现持续增加趋势, 并且在鳞茎膨大期, MDA含量的增加幅度最大, 这可能是该时期百合植株处于生长旺盛期, 从而向土壤中分泌了更多的化感自毒物质, 对植株细胞膜的完整性破坏程度加强, 并且随连作年限的延长破坏程度加大, 最终导致MDA含量大幅度增加。
 | 图5 连作对百合不同生育时期叶片MDA含量的影响Fig.5 Effect of continuous cropping on MDA content in leaves of lily at different growth stages |
采用气质联用色谱仪(GC-MS)分析不同处理土壤中潜在的化感自毒物质。结果发现, 正茬土壤中存在9种化合物, 主要包括:间苯二甲酸二辛酯及1, 3-二甲苯等。连作2年土壤中存在15种化合物, 主要包括:对苯二甲酸二辛酯、甲苯、2, 3-丁二醇和2, 2'-亚甲基双-(4-甲基-6-叔丁基苯酚)等成分。连作4年土壤中共鉴定出17种化合物, 主要包括:对苯二甲酸二辛酯、对二甲苯和2, 2'-亚甲基双-(4-甲基-6-叔丁基苯酚) 等物质。3处理土壤中相对含量最高的成分均为酯类化合物。其中, 正茬土壤中间苯二甲酸二辛酯相对含量高达80.73%, 连作2年和连作4年土壤中相对含量最高的组分均为对苯二甲酸二辛酯, 分别为27.47%和41.43%。表3、表4和表5中列出的化感物质均属GC-MS分析匹配度大于90%的化合物。
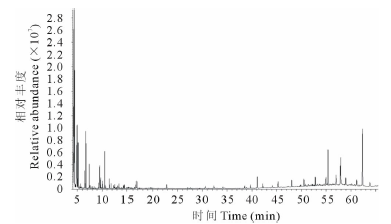 | 图6 正茬土壤萃取液GC-MS色谱图Fig.6 GC-MS chromatogram of the stubble soil extraction |
 | 图7 连作2年土壤萃取液GC-MS色谱图Fig.7 GC-MS chromatogram of the continuous cropping soil extraction for 2 years |
 | 图8 连作4年土壤萃取液GC-MS色谱图Fig.8 GC-MS chromatogram of the continuous cropping soil extraction for 4 years |
| 表3 正茬土壤中存在的主要化感成分 Table 3 The main allelochemicals in the stubble soil |
| 表4 连作2年土壤中存在的主要化感成分 Table 4 The main allelochemicals in the continuous cropping soil for 2 years |
| 表5 连作4年土壤中存在的主要化感成分 Table 5 The main allelochemicals in the continuous cropping soil for 4 years |
杜家方等[15]研究发现, 在地黄(Rehmannia glutinosa)整个生育期内, 重茬地黄的各项形态指标均小于正茬地黄。康亚龙等[16]认为, 加工番茄(Lycopersicon esculentum)连作3年后, 在其整个生育进程中均表现出连作障碍效应且随连作年限的延长呈显著增强趋势。笔者对正茬和连作百合进行对比研究发现, 连作能够抑制百合植株的生长, 抑制作用始于苗期而贯穿整个生育期, 并且, 随连作年限的延长作用程度增强。试验结果与上述研究结论相一致。
本研究还显示, 整个生育期, 连作处理中鳞茎的生长一直受到抑制, 并且于成熟期发现, 与正茬相比, 连作条件下鳞茎个头较小, 重量较轻, 且表皮颜色发黄, 有腐烂现象, 这可能就是导致百合产量下降的主要原因。前人[15]研究结果显示作物产量的高低直接受到地上部分光合产物及根系吸收养分和水分的影响, 在本试验中主要是基于百合地上部分生理特性的变化来研究兰州百合对连作的响应机制。然而, 根系是矿物养分和水分的吸收器官, 连作对百合根系生长的阻碍效应必将影响根系的正常功能, 造成鳞茎及地上部分营养及水分胁迫, 导致植株生长缓慢, 最终影响百合产量。但是, 关于连作百合根系生理变化方面的研究未见报道, 其作用机制有待我们进一步研究。
本试验结果显示, 连作降低了百合叶片中净光合速率(Pn), 从而降低了植株的光合作用能力。研究认为[17, 18]光合速率的下降或是由于气孔关闭导致的CO2供应不足即气孔因素引起, 或是叶肉细胞光合能力受害所致的叶绿素含量下降等非气孔因素限制, 或者是两种因素兼而有之。而Pn的下降受气孔因素还是非气孔因素限制则取决于Pn、Gs和Ci的变化方向, 若Pn、Gs和Ci均下降, 说明Pn的下降受气孔因素限制, 若Pn和Gs下降, Ci上升, 表明Pn的下降受非气孔因素限制[19, 20]。本试验T1中Pn、Gs分别下降28.03%和14.25%, T2中分别下降40.59%和23.97%, 两处理Ci则分别增加了18.00%和21.20%, 叶片中Chl总量随连作年限的增加呈降低趋势, 说明百合植株Pn的下降是受非气孔因素限制。Chl总量下降引起Pn的降低, 加之叶绿体的捕光能力及能量传递速率受阻, 导致光合能力下降, 影响植株的正常生长, 最终导致百合产量下降。
正常情况下, 植物细胞内抗氧化酶系统能够维持活性氧的产生和清除, 但在逆境环境中, 活性氧的代谢平衡遭到破坏, 细胞膜脂过氧化程度升高, MDA含量随之增加[21]。本研究表明, 随着连作年限的增加, 叶片中CAT、SOD活性下降越明显, 引起细胞内活性氧的大量累积, 从而激发了POD活性, 使其含量增加, 提高植物抗胁迫能力。随着连作时间的延长, CAT、SOD活性先升高后降低, POD活性持续下降, 细胞膜脂过氧化水平不断提高, MDA含量持续增加。表明连作可以影响百合植株的防御系统和次生代谢过程, 使百合植株抗氧化系统紊乱, 阻碍次生代谢, 引起植株衰老, 导致产量和品质下降。
研究显示[22, 23]自毒物质多数来源于植物次生代谢产物, 而次生代谢产物的合成与累积往往受制于所处环境的变化, 环境胁迫下可提高次生代谢产物的含量。本试验结果显示, 连作2年和4年土壤中化感物质种类较正茬明显增加, 且主要物质对苯二甲酸二辛酯在连作4年土壤中相对含量达到41.43%, 较2年土壤增加了19.77%, 表明随着连作年限的增加, 环境胁迫强度加大, 导致百合根际环境中化感物质种类和数量增加。反之, 自毒物质的累积又是引起连作障碍的主要因素之一[24, 25]。吴秀华等[26]认为巨桉(Eucalyptus grandis)根系中存在的自毒物质能够破坏光合结构及光合生理特性, 降低光合色素含量, 从而影响巨桉生物量的积累。张如义等[27]也认为核桃(Juglans regia)叶内含有的化感物质能够影响幼苗抗氧化酶系统, 破坏活性氧代谢平衡, 导致膜脂过氧化, 最终抑制幼苗的生长。本试验也证实了植前连作土壤中残存的自毒成分是降低百合植株光合能力, 破坏植株体内抗氧化酶活性, 抑制自身生长, 导致障碍效应发生的因素之一。同时, 在生长发育过程中, 百合植株又进一步产生多种化感物质, 通过挥发、雨淋等方式对同茬及下茬百合产生一定的障碍效应。
综上所述, 连作能够抑制百合植株的生长, 一方面, 通过降低百合植株叶绿素含量, 影响净光合速率, 从而降低光合作用能力。另一方面, 通过抑制抗氧化酶活性, 造成活性氧代谢失调, 膜脂过氧化程度升高, 破坏细胞膜完整性, 从而抑制百合植株生长, 阻碍鳞茎膨大, 引起百合产量和品质下降。连作土壤中存在的各种化感物质是影响百合植株生理变化, 导致障碍效应发生的主要原因之一。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|