
{kind=link}
{kind=link}
{kind=link}
不同生态恢复措施下宁夏黄土丘陵区典型草原植物群落数量分类和演替
[贾希洋2  , 马红彬
, 马红彬1, 2, * , 周瑶2 , 张蕊2 , 宿婷婷2 , 张双乔2 , 张俊2 ]
, 马红彬, 周瑶|
|


作者简介:贾希洋(1997-),男,甘肃永昌人,在读硕士。E-mail:573342187@qq.com
进行植物群落数量分类和演替研究对退化草地生态恢复具有重要意义。以宁夏黄土丘陵区典型草原为对象,借助植被数量生态学方法,对封育、水平沟和鱼鳞坑3种生态恢复措施实施0(1年)、3、6、10和15年下的草地植物群落进行了数量分类,研究了群落演替序列及其与土壤因子的关系。结果表明:(1)应用双向指示种(TWINSPAN)结合除趋势对应分析(DCA),将研究样地植物群落类型划分为8个。(2)封育0~15年,植被演替序列为本氏针茅+百里香群落→本氏针茅+大针茅群落→百里香+本氏针茅群落→本氏针茅+大针茅群落→本氏针茅+铁杆蒿群落;水平沟和鱼鳞坑整地后1年到15年,植被演替序列分别为沙打旺+白草群落→早熟禾+赖草群落→本氏针茅+百里香群落→百里香+赖草群落→本氏针茅+大针茅群,沙打旺+白草群落→早熟禾+本氏针茅群落→本氏针茅+百里香群落→百里香+本氏针茅群落;随着恢复年限增加,鱼鳞坑和水平沟措施下植物优势物种变化总趋势是根茎型—疏丛型—密丛型植物,但也存在非线性或螺旋式的变化; 3种恢复措施下,植物群落盖度和地上生物量随恢复年限增加呈上升趋势,其中封育草地增加速度最快,鱼鳞坑最慢。(3)除趋势典范对应分析(DCCA)表明,土壤容重、有机质、真菌、全磷和蛋白酶是影响该区植物群落分布的主要土壤因子。
, MA Hong-bin, ZHOU YaoRestoration measures strongly influence vegetation communities. To design effective restoration measures for the Loess Hilly Region, it is important to classify the flora and study the succession of degraded grassland plant communities on the typical steppe in Ningxia. In this study, we explored the relationship between the plant succession process and soil factors under three ecological restoration measures (enclosure, contour trench, and fish-scale pits) implemented for 0 (1), 3, 6, 10 and 15 years by using Two Indicators Species Analysis (TWINSPAN) combined with a detrended correspondence analysis (DCA). The main results were as follows: (1) According to the TWINSPAN-DCA, the vegetation in the study plot was divided into eight communities. (2) With increasing exclosure time, the vegetation succession sequence was as follows: Stipa bungeana+Thymus mongolicus communities→ S. bungeana+Stipa grandis communities→ T. mongolicus+S. bungeana communities→ S. bungeana+S. grandis communities→ S. bungeana+Artemisia sacrorum communities. The succession process from 1 to 15 years in the contour trench grassland was as follows: Astragalus adsurgens+Pennisetum flaccidum communities→ Poa annua+Leymus secalinus communities→ S. bungeana+T. mongolicus communities→ T. mongolicus+L. secalinus communities→ S. bungeana+S. grandis communities. The succession process from 1 to 15 years in the fish-scale pits grassland was as follows: A. adsurgens+P. flaccidum communities→ P. annua+S. bungeana communities→ S. bungeana+T. mongolicus communities→ T. mongolicus+S. bungeana communities. In the contour trench grassland and the fish-scale pits grassland, the trend in the dominant vegetation type over time was rhizome type-sparse type-dense cluster type, but there were also non-linear or spiral changes. The coverage of the plant community and aboveground biomass increased over time under all three restoration measures, and the growth rate was fastest in enclosed grassland and slowest in the fish-scale pits grassland. (3) The results of a detrended canonical correspondence analysis indicated that soil bulk density, organic matter, fungi, total phosphorus, and protease activity are the main soil factors affecting the vegetation communities in this area.
群落特征是植被恢复与重建的基础[1]。在全球草地退化的背景下, 退化草地生态系统的植被变化引起了广泛的关注[2, 3, 4]。受长期自然因素和人为因素的影响, 黄土高原丘陵区的天然植被退化严重[5]。随着国家生态环境治理工程的相继实施, 黄土丘陵区一些地区在草原生态恢复中实施了围栏封育、开挖水平沟和鱼鳞坑等工程措施来促进天然植被的恢复[6]。目前宁夏黄土丘陵区典型草原已全部实施了封育禁牧措施, 水平沟、鱼鳞坑地表扰动面积已占到整个丘陵草原面积的20%以上, 这些恢复措施使原有的天然草地植物群落特征及物种多样性发生了改变[7, 8]。研究表明, 典型草原植被盖度、地上生物量、丰富度和多样性随封育年限增加呈先上升后下降变化[9, 10], 主要植物种群空间分布格局在封育、水平沟和鱼鳞坑措施下变化显著[11]。草地植被恢复过程中土壤种子库与地上植被存在一定关系[12, 13]、优势植物生态位特征发生了变化[14]。
随着数量生态学的发展, 数量分类和排序已成为认识一定区域内植被分布格局特征的重要手段, 它们在群落分析中的使用可进一步揭示植物种、植物群落与环境间的生态关系, 被广泛应用于植被群落研究。采用数量分类和排序, 一些学者对植物群落类型进行了划分, 探讨了群落分布和环境因子的关系[15], 分析了植被演替规律[16, 17]。植物群落数量分类目前应用最普遍的方法有双向指示种(TWINSPAN)、除趋势对应分析(DCA)、除趋势典范对应分析(DCCA)、典范对应分析(CCA)等, 这些方法为更加客观、准确地反映植被与环境之间的生态关系提供了有效途径[18]。目前有关黄土丘陵区典型草原植被特征已有较多研究, 但有关生态恢复措施下植物群落数量分类和演替的报道较少。因此, 应用TWINSPAN、DCA和DCCA, 对宁夏黄土丘陵区典型草原封育、鱼鳞坑和水平沟3种不同恢复措施下的植物群落进行数量分类与排序、探讨生态恢复过程中植物群落演替状况、分析群落与土壤因子间的关系, 以期为当地生态建设与恢复提供依据。
试验区位于宁夏南部云雾山草原国家自然保护区, 介于东经106° 21'-106° 27', 北纬36° 10'-36° 17'。海拔1800~2150 m之间, 为典型的黄土低山丘陵区, 是中国保护较好的典型草原之一。属于典型的中温带大陆性气候, 年均气温5 ℃, ≥ 0 ℃的年积温2882 ℃, 年无霜期137 d, 年降水量445 mm。地带性土壤主要为山地灰褐土, pH在8.1左右。地带性植被为典型草原, 主要分布有本氏针茅(Stipa bungeana)、大针茅(Stipa grandis)、百里香(Thymus mongolicus)、阿尔泰狗哇花(Heteropappus altaicus)、猪毛蒿(Artemisia scoparia)、铁杆蒿(Artemisia sacrorum)、赖草(Leymus secalinus)、白草(Pennisetum flaccidum)、猪毛菜(Salsola collina)、星毛委陵菜(Potentilla acaulis)等。多年的生态建设使该区天然草原中分布有不同封育年限的草地、大量的水平沟和鱼鳞坑, 其中水平沟是在天然草地上沿等高线隔带设置, 上下水平沟带间距14 m, 沟宽1 m, 上埂高0.6 m, 下埂高0.2 m; 鱼鳞坑是在天然草地上呈品字形设置, 坑距3 m, 下埂弧长1.5 m左右, 上埂高0.6 m, 下埂高0.2 m。当地水平沟和鱼鳞坑整地时一般将表土回填, 回填深度约0.4 m, 整地当年在沟(坑)中种植沙打旺(Astragalus adsurgens)。
在试验区海拔、坡度和坡向尽量接近的地段, 选择不同年限水平沟、鱼鳞坑、封育草地和未封育草地(放牧草地)为研究对象, 设置未封育、封育3年(禁牧封育3年)、6、10和15年, 水平沟1年(水平沟整地1年后)、3、6、10和15年, 以及鱼鳞坑1年(鱼鳞坑整地1年后)、3、6、10和15年草地共15个处理, 每个处理3次重复。试验样地概况见表1。
| 表1 试验样地概况 Table 1 Details of the test sample |
1.3.1 植被调查 于2015和2016年8月, 观测各个样地植物组成, 分物种测定高度、密度、频度、盖度及地上生物量。样方面积为1 m× 1 m, 3次重复。其中高度为自然高度, 对每个物种随机测定30株, 求其平均高度; 密度采用统计单位面积株数法测定, 频度则用样圆法来测定, 盖度采用针刺法测定, 地上生物量测定时分种齐地刈割样方内植物, 带回室内65 ℃烘至其恒重。
1.3.2 土壤取样 测定植被的同时, 在各样地采用“ S” 形五点法采集土样。采集时除去凋落物, 用土钻按0~10 cm, 10~20 cm, 20~30 cm, 30~40 cm土层取样, 3次重复; 用环刀法测定土壤容重[19], 3次重复。将采集到土样装在塑封袋放入4 ℃的保温箱中带回实验室。在室内去除凋落物和根后, 过2 mm筛后将土样分为两份, 一份保存在4 ℃下用于土壤微生物数量和土壤酶活性的测定; 另一份常温存放用于土壤颗粒组成和养分测定。
1.4.1 土壤颗粒组成测定 用Microtrac S3500激光粒度分析仪测定土壤颗粒组成[20]。
1.4.2 可培养土壤微生物数量的测定 可培养微生物群落的数量选真菌、细菌和放线菌代表土壤总体微生物数量。采用平板涂布法测定土壤细菌(稀释梯度为10-2、10-3、10-4)、放线菌( 稀释梯度为10-3、10-4、10-5)和真菌(稀释梯度为100、10-1、10-2)数量, 每个稀释梯度3次重复, 其中细菌采用牛肉膏蛋白胨琼脂培养基, 真菌采用孟加拉红培养基, 放线菌采用高氏1号培养基[21]。
1.4.3 土壤酶活性测定 蔗糖酶用3, 5-二硝基水杨酸比色法, 蛋白酶用福林酚法, 过氧化氢酶活性采用高锰酸钾滴定法测定, 磷酸酶采用磷酸苯二钠比色法, 脲酶采用靛酚比色法[22]。
1.4.4 土壤养分的测定 土壤有机质含量测定采用重铬酸钾容量法, 土壤全氮含量测定采用全自动凯氏定氮法测定, 土壤速效钾含量测定采用近红外光谱法, 土壤速效氮采用碱解扩散法, 土壤全磷含量采用氢氧化钠碱熔-钼锑抗比色法测定[23, 24]。
2.1.1 群落类型TWINSPAN分类 对15个处理49种植物进行数量分类, 图1是TWINSPAN分类结果的树状图, 方框内的代号代表各处理编号, 方框下数字表示的是群落编号。第一次划分的对象为处理总数, 以沙打旺(+)作为区别种, 将其划分为两部分, 包括沙打旺+白草群落(2个处理)和其他13个处理。第二次划分的对象为第一次划分出来的13个处理, 以大针茅(+)为指示种, 将其划分为两部分, 分别包括7个处理和6个处理。第三次划分的对象为第二次划分出来的7个处理, 以猪毛蒿(+)为区分种, 将其划分为两部分, 分别包括6个处理和早熟禾(Poa annua)+赖草群落。依次类推, 依据TWINSPAN分类结果的指示种, 最终将这15个处理样地划分为8类群落。每个群落的主要特征如下:
Ⅰ .沙打旺+白草群落
包括S1、Y1处理, 优势种为沙打旺, 次优势种为白草, 伴生有百里香、冰草(Agropyron cristatum)、冷蒿(Artemisia frigida)、阿尔泰狗哇花、猪毛菜、秦艽(Gentiana macrophylla)、西藏点地梅(Androsace mariae)、山苦荬(Ixeris denticulata)、狼毒(Stellera chamaejasme)等。群落高度在3.09~9.33 cm, 盖度在18.76%~25.33%, 地上生物量在21.14~29.94 g· m-2。
Ⅱ .早熟禾+赖草群落
包括S3处理, 优势种为早熟禾, 次优势种为赖草, 伴生有本氏针茅、阿尔泰狗哇花、星毛委陵菜、米口袋(Gueldenstaedtia verna)、风毛菊(Saussurea japonica)、扁蓿豆(Melissitus ruthenica)等。群落高度在11.65~41.86 cm, 盖度在69.01%~74.34%, 地上生物量在36.99~47.27 g· m-2。
Ⅲ .百里香+赖草群落
包括S10处理, 优势种为百里香, 次优势种为赖草, 伴生有本氏针茅、猪毛蒿、狼毒、黄花棘豆(Oxytropis ochrocephala)、西藏点地梅、山苦荬、糙隐子草(Cleistogenes squarrosa)等。群落高度在2.92~13.78 cm, 盖度在60.34%~72.18%, 地上生物量在52.89~77.50 g· m-2。
Ⅳ .本氏针茅+百里香群落
包括F0、S6、Y6和Y10处理, 优势种为本氏针茅, 次优势种为百里香, 伴生有赖草、狼毒、早熟禾、猪毛蒿、阿尔泰狗哇花、西藏点地梅、独行菜(Lepidium apetalum)、风毛菊等。群落高度在3.30~18.41 cm, 盖度在40.23%~77.65%, 地上生物量在27.68~73.06 g· m-2。
Ⅴ .早熟禾+本氏针茅群落
包括Y3处理, 优势种为早熟禾, 次优势种为本氏针茅, 伴生有铁杆蒿、猪毛蒿、阿尔泰狗哇花、白草、狼紫草(Lycopsis orientalis)、星毛委陵菜等。群落高度在2.80~20.94 cm, 盖度在24.22%~27.12%, 地上生物量在60.93~76.65 g· m-2。
Ⅵ .本氏针茅+大针茅群落
包括F3、F10和S15处理, 优势种为本氏针茅, 次优势种为大针茅, 伴生有赖草、百里香、猪毛蒿、阿尔泰狗哇花、披针叶黄华(Thermopsis lanceolata)、打碗花(Calystegia hederacea)、蒙古芯芭(Cymbaria mongolica)等。群落高度在4.21~23.94 cm, 盖度在67.16%~83.97%, 地上生物量在54.41~98.93 g· m-2。
Ⅶ .百里香+本氏针茅群落
包括F6、Y15处理, 优势种为百里香, 次优势种为本氏针茅, 伴生有赖草、铁杆蒿、早熟禾、香青兰(Dracocephalum moldavica)、裂叶堇菜(Viola dissecta)、独行菜等。群落高度在8.31~35.37 cm, 盖度在37.82%~73.41%, 地上生物量在54.66~82.07 g· m-2。
Ⅷ .本氏针茅+铁杆蒿群落
包括F15处理, 优势种为本氏针茅, 次优势种为铁杆蒿, 伴生有百里香、大针茅、赖草、二裂委陵菜(Potentilla bifurca)、裂叶堇菜、远志(Polygala tenuifolia)、柴胡(Bupleurum chinense)、白草等。群落高度在4.12~34.96 cm, 盖度在67.93%~88.07%, 地上生物量在93.58~116.61 g· m-2。
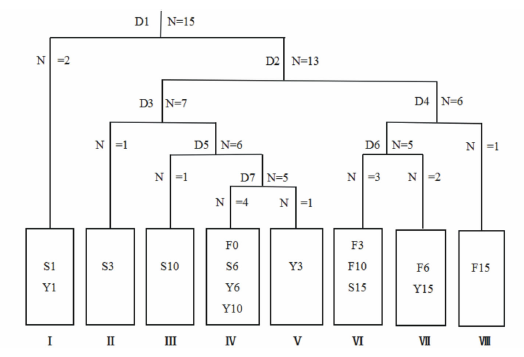 | 图1 不同恢复措施下典型草原植物群落TWINSPAN分类结果树状示意图Fig.1 Different restoration measures under the typical grassland plant community TWINSPAN classification results tree schematic diagram |
2.1.2 植物群落DCA排序 TWINSPAN分类可能会产生边界样地, 甚至有时候也会错分样地, 更加准确的划分结果应该是TWINSPAN分类结合DCA排序[27, 28], 首先用物种数据进行DCA分析, 在4个排序轴中梯度长度最大值3.215, 因此选择单峰模型和线性模型均合适, 论文选择了单峰模型(DCA)。采用DCA对15个处理内49个植物种进行排序, 结果显示将样方划分成8个区(图2), DCA排序结果与TWINSPAN分类结果较吻合, 验证了TWINSPAN分类结果的合理性。
根据TWINSPAN分类(图1)和DCA排序结果(图2), 确定了不同恢复措施植物群落演替序列。
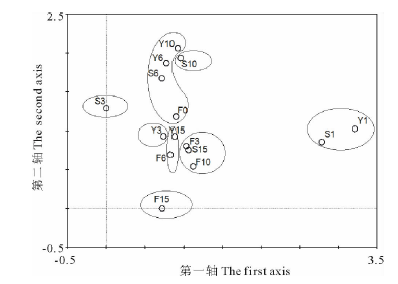 | 图2 不同恢复措施下15个处理植物群落DCA排序Fig.2 Different restoration measures under the 15 treatment of plant community DCA ordination |
2.2.1 封育草地植被演替序列 自封育0年到15年, 植被演替序列为:本氏针茅+百里香群落(F0)→ 本氏针茅+大针茅群落(F3)→ 百里香+本氏针茅群落(F6)→ 本氏针茅+大针茅群落(F10)→ 本氏针茅+铁杆蒿群落(F15)。结合前面的群落特征可知, 随着恢复年限的延长, 植物优势物种变化不大, 群落高度以放牧草地最低, 其他封育年限较为接近, 盖度和地上生物量虽有波动但总体呈现上升趋势。
2.2.2 水平沟植被演替序列 水平沟1年到15年, 植被演替序列为:沙打旺+白草群落(S1)→ 早熟禾+赖草群落(S3)→ 本氏针茅+百里香群落(S6)→ 百里香+赖草群落(S10)→ 本氏针茅+大针茅群落(S15)。可见, 整地后的1年, 植物群落优势种为整地后补播的沙打旺和根茎禾草, 随着水平沟整地年限的延长, 优势物种逐渐演替为丛生禾草, 到整地后15年, 优势种与封育6年草地一致。群落高度随着整地后时间的延迟无明显变化规律, 盖度和地上生物量呈上升趋势。
2.2.3 鱼鳞坑植被演替序列 鱼鳞坑1年到15年, 植被演替序列为:沙打旺+白草群落(Y1)→ 早熟禾+本氏针茅群落(Y3)→ 本氏针茅+百里香群落(Y6、Y10)→ 百里香+本氏针茅群落(Y15)。鱼鳞坑整地后1年, 植物优势种以补播的沙打旺和根茎禾草— 白草为主, 与水平沟整地后1年一致。随着恢复年限延长, 优势物种逐渐演替为本氏针茅、百里香, 到整地后15年, 优势种与封育10年草地一致。群落高度随着恢复时间的延长无明显变化规律, 但盖度和地上生物量呈增加趋势。
土壤因子对植物群落的组成、分布及演替有很大的影响, 为进一步分析植物群落和土壤环境因子的关系, 对15个处理样地与15个土壤环境因子采用除趋势典范对应分析(DCCA)。前两排序轴的特征值之和占全部排序轴总特征值的71.6%, 解释了群落分布与各个土壤因子间关系的大部分信息, 排序结果较为理想。排序结果表明(表2), 土壤蛋白酶和全磷与第一排序轴的相关性最大, 土壤容重、有机质和真菌与第二排序轴的相关性最大, 第一排序轴主要反映了土壤蛋白酶和全磷的变化, 自左向右土壤蛋白酶和全磷均降低, 第二排序轴主要反映土壤容重、土壤有机质和真菌的变化, 从上至下, 土壤容重降低、有机质和真菌升高。可见, 土壤容重、有机质、真菌、全磷和土壤蛋白酶是影响该区群落分布的主要土壤因子。
DCCA排序图中, 箭头所在象限代表环境因子与各个排序轴之间正负相关性, 其连线的长度表示环境因子和物种分布的相关程度, 连线越长, 表明相关性越大, 反之越小; 箭头连线与排序轴的夹角则表示该环境因子与排序轴的相关性, 其夹角越小, 说明该环境因子与排序轴的相关性越大[29]。不同生态恢复措施下植物群落在排序图第一、二轴的相关位置, 反映了典型草原土壤环境因子对其植物群落空间分布的影响(图3)。从图3可见, 随着封育年限的增加, 除土壤容重和过氧化氢酶活性呈现逐渐减小外, 其他土壤环境因子总体上呈现增加变化。水平沟和鱼鳞坑措施下, 从1年到15年, 土壤环境因子无明显变化规律。从各个土壤因子箭头连线的长短可以看出, 15个土壤因子对典型草原群落的分布存在不同程度的影响。将15个处理垂直投影在环境因子向量延长线上, 再比较到各个环境因子的距离, 距离越远说明影响程度越低, 反之, 则影响程度越大。可以看出S1、Y1和S3三个植物群落主要生长在土壤紧实、黏粒含量少、氮磷钾和有机质含量较低的生境。F15、F10、F3、S15、F6、Y15、Y3、F0植物群落主要分布于土壤疏松、土壤黏粒含量多、氮磷钾和有机质含量较高的生境。
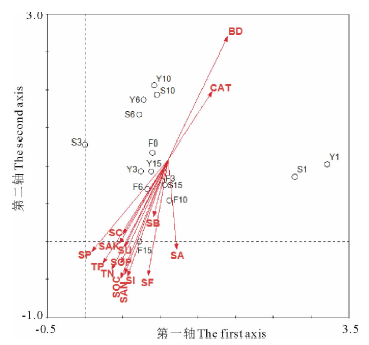 | 图3 不同群落与土壤环境因子的DCCA排序 SC:土壤黏粒; BD:土壤容重; SOC:土壤有机质; TN:土壤全氮; TP:土壤全磷; SAK:土壤速效钾; SAN:土壤速效氮; SF:真菌; SB:细菌; SA:放线菌; SU:脲酶; SP:蛋白酶; SOP:磷酸酶; SI:蔗糖酶; CAT:过氧化氢酶。Fig.3 Different communities and soil environmental factors in DCCA ordination diagram SC: Soil clay; BD: Soil bulk density; SOC: Soil organic matter; TN: Soil total nitrogen; TP: Soil total phosphorus; SAK: Soil available potassium; SAN: Soil available nitrogen; SF:Fungi; SB: Bacteria; SA: Actinomycetes; SU: Urease; SP: Protease; SOP: Phosphatase; SI: Sucrase; CAT: Catalase. |
| 表2 土壤环境因子与DCCA排序轴的相关系数 Table 2 Soil environment factor and sort DCCA axis of the phase relationship |
应用TWINSPAN分类结合DCA排序, 将研究样地植物群落类型划分为8个群落, 各个植物群落在植物组成、盖度和地上生物量等方面存在一定差异。沙打旺+白草群落的高度、盖度及地上生物量在所有群落中最低, 与其整地后恢复年限较短有关。TWINSPAN分类和DCA排序二者结合更能客观准确划分群落类型, 两种方法可互相对比验证。本研究中WINSPAN分类和DCA排序结果的一致性也说明了这两种方法对宁夏黄土丘陵区典型草原植被群落数量分析结果的可靠性, 与陶楚等[30]和代雪玲等[31]同时使用上述两种方法来进行群落数量分类的研究结果一致。另外, 在具体分类过程中, 还应该充分考虑群落生境特征和各个物种在群落内的生态位, 才能获得更符合实际的分类结果。
长期定位研究可获得较为系统的植被演替序列, 但由于实践中永久研究样地建立受诸多因素限制, 同一样地时间序列上的植被变化在研究实践中往往较难获得, 空间梯度代替时间梯度的方法是学者普遍认可的研究群落演替的方法[32, 33]。不同生态恢复措施会使土壤性状发生变化, 进而影响地上植被[34]。鱼鳞坑和水平沟整地增加了土壤透气性, 人为又补播了沙打旺, 白草为根茎型植物, 疏松的土壤更有利于其根系的生长。因此水平沟和鱼鳞坑整地1年时沙打旺为优势种, 白草为次优势种。随着演替的进行, 土壤有机质和土壤养分逐渐增加[35], 土壤黏粒含量亦会增大, 土壤透气性逐渐下降[36], 群落逐步演替成以早熟禾(疏丛型)为优势种、以针茅(密丛型)为优势种的植物群落。总体上, 论文基于分类和排序结果确定的封育、水平沟和鱼鳞坑3种恢复措施下草地植被的演替序列符合典型草原植被演替规律。但是, 封育3年和10年草地群落优势种相同、鱼鳞坑6年与10年草地群落优势种相同, 说明植被演替有时会呈现非线性或螺旋式的变化[37], 这种变化也与鱼鳞坑和水平沟整地时一些植物有生命力的根系被填埋到浅层土壤中, 在适宜条件下有的会再次萌发成新的植株、甚至演替成群落优势种有关。就植物物种组成、盖度和地上生物量而言, 3种措施下, 封育草地增加速度最快, 水平沟次之, 鱼鳞坑最慢。
随着封育年限的增加, 凋落物逐渐积累, 土壤有机质、微生物数量和大部分酶活性含量逐渐增加, 但土壤容重与过氧化氢酶活性呈逐渐减小, 与大部分学者研究结果一致[38, 39, 40]。整地后1年到15年水平沟和鱼鳞坑土壤环境因子变化规律不明显与整地时将先前的地上植物和地下根系回填到土壤中的分解状况、整地后播种的沙打旺的短期生长以及整地后地上植被的自然恢复等因素有关。由于CCA存在“ 弓形效应” , DCCA可以克服这一缺点, 大多数学者认为DCCA的结果优于CCA[41, 42], 本研究用DCCA排序探讨了黄土丘陵区典型草原植物群落与环境因子之间的关系, 发现土壤环境因子的空间分布较好地揭示了群落的分布, 这与多数研究结果相符[43, 44]。DCCA排序只是在半定量的层面上解释植物群落与环境因子之间的关联, 若要全面了解植物群落与环境之间的定量关系, 除土壤因子外, 地形、气候和植物种内种间作用等都会影响群落的分布格局[45], 运用数学模型进行模拟和定量化解释会更加全面了解植物群落与环境之间的定量关系[46], 这还有待于进一步研究。
(1)应用TWINSPAN分类结果结合DCA排序, 将黄土丘陵区典型草原封育、水平沟和鱼鳞坑恢复措施下0(1)-15年植物群落类型划分沙打旺+白草、早熟禾+赖草、百里香+赖草、本氏针茅+百里香、早熟禾+本氏针茅、本氏针茅+大针茅、百里香+本氏针茅、本氏针茅+铁杆蒿8个群落。
(2)草地封育0年到15年, 植被演替序列为本氏针茅+百里香群落→ 本氏针茅+大针茅群落→ 百里香+本氏针茅群落→ 本氏针茅+大针茅群落→ 本氏针茅+铁杆蒿群落; 水平沟和鱼鳞坑整地后1年到15年, 植被演替序列分别为沙打旺+白草群落→ 早熟禾+赖草群落→ 本氏针茅+百里香群落→ 百里香+赖草群落→ 本氏针茅+大针茅群落, 沙打旺+白草群落→ 早熟禾+本氏针茅群落→ 本氏针茅+百里香群落→ 百里香+本氏针茅群落。3种恢复措施下, 群落盖度、地上生物量随恢复年限增加呈增加趋势, 其中封育草地增加最快, 水平沟次之, 鱼鳞坑最慢。
(3)除趋势典范对应分析(DCCA)表明, 土壤容重、有机质、真菌、全磷和土壤蛋白酶是影响该区植物群落分布的主要土壤因子。整地1年后的鱼鳞坑、整地1年和3年后的水平沟其植物群落可生长在土壤紧实、黏粒含量少、氮磷钾和有机质含量较低的生境, 封育草地和其他年限较长的水平沟、鱼鳞坑植物群落分布于土壤疏松、黏粒含量多、氮磷钾和有机质含量较高的生境。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|