


{kind=link}
{kind=link}
{kind=link}
过氧化氢提高燕麦幼苗耐碱性的活性氧代谢和渗透调节
[刘建新*  , 欧晓彬, 王金成, 刘秀丽]
, 欧晓彬, 王金成, 刘秀丽]
, 欧晓彬, 王金成, 刘秀丽]
|
|


作者简介:刘建新(1964-),男,甘肃通渭人,教授,本科。E-mail: liujx1964@163.com
为探明过氧化氢(H2O2)提高燕麦耐碱性的作用,以‘定莜6号’幼苗为材料,采用珍珠岩栽培方法,在幼苗三叶一心期根部浇灌75 mmol·L-1 NaHCO3或添加二甲基硫脲(DMTU,H2O2淬灭剂)或抗坏血酸(ASA,H2O2清除剂)模拟碱胁迫,通过叶面喷施0.01 mmol·L-1 H2O2来观测H2O2对碱胁迫下幼苗生长、活性氧代谢和渗透溶质积累的影响。结果表明:喷施H2O2 能够缓解NaHCO3胁迫对燕麦幼苗生长的抑制,降低幼苗叶片
Hydrogen peroxide (H2O2) is a signaling molecule with diverse physiological functions in plants. The objective of this study was to determine whether exogenous H2O2 could improve the alkali tolerance of oat ( Avena nuda) seedlings. Seedlings of oat cv. Dingyou No.6 at the three-leaf stage were irrigated with 75 mmol·L-1 NaHCO3 solution containing dimethylthiourea (DMTU, a quencher of H2O2) or ascorbate (ASA, a scavenger of H2O2), and the leaves were sprayed with 0.01 mmol·L-1 H2O2. Seedlings irrigated with water served as the control. After 7 days of treatment, seedling growth, chlorophyll content, active oxygen metabolism, and osmotic accumulation in leaves were measured. The results showed that treatments with exogenous H2O2 remarkably relieved the growth inhibition caused by NaHCO3 stress, and significantly decreased the contents of
土壤盐碱化是制约农业发展的重要因素之一。中国盐碱土面积3.467× 107 hm2(不包括滨海滩涂), 其中碱土面积为8.667× 105 h
过氧化氢(hydrogen peroxide, H2O2)是植物代谢过程中产生的具有毒害作用的活性氧, 但新近研究发现, 低浓度H2O2作为信号分子参与植物对盐碱胁迫适应的调控[6]。研究表明, H2O2作为根源信号介导盐胁迫诱导的蚕豆(Vicia faba)气孔关闭[7]; 外源H2O2能够缓解氧化损伤和促进胁迫蛋白表达增强小麦(Triticum aestivum)耐盐性[8], 并减轻受旱黄瓜(Cucumis sativus)叶绿体膜的伤害[9], 提高葡萄(Vitis)抗冷性[10]和水稻(Oryza sativa)镉胁迫耐性[11]。外源H2O2是否参与碱胁迫燕麦生理响应调控, 从而提高燕麦耐碱性, 目前研究未见报道。本试验以甘肃省广泛种植的‘ 定莜6号’ 燕麦新品种为材料, 研究喷施H2O2对NaHCO3胁迫下幼苗生长及叶片活性氧代谢和渗透调节的影响, 探讨H2O2提高燕麦耐碱性的作用及其生理机制, 以期为燕麦耐盐碱栽培提供理论依据。
试验于2015年4-7月在甘肃省高校陇东生物资源保护与利用省级重点实验室日光温室进行。供试燕麦品种 ‘ 定莜6号’ 种子用1%次氯酸钠表面消毒后在瓷盘中25 ℃催芽, 选露白一致的萌发苗播种在装满珍珠岩的塑料盆(口径20 cm, 高14 cm)中, 浇水后置日光温室培养, 昼/夜温度(23± 6) ℃/(17± 7) ℃, 相对湿度60%~80%, 光照强度360~710 μ mol· m-2· s-1。幼苗两叶一心期选留长势一致的壮苗约200株/盆, 三叶一心期用Hoagland营养液浇灌一次, 然后进行处理。
试验设6个处理:1) CK(对照):喷蒸馏水+根施蒸馏水; 2) T1:喷蒸馏水+根施75 mmol· L-1 NaHCO3; 3) T2:喷0.01 mmol· L-1 H2O2+根施75 mmol· L-1 NaHCO3; 4) T3:喷0.01 mmol· L-1 H2O2+根施蒸馏水; 5) T4:喷 0.01 mmol· L-1 H2O2+根施75 mmol· L-1 NaHCO3+根施5 mmol· L-1二甲基硫脲(DMTU); 6) T5:喷0.01 mmol· L-1 H2O2+根施75 mmol· L-1 NaHCO3+根施2 mmol· L-1 抗坏血酸(ASA)。
DMTU是H2O2的淬灭剂, ASA是H2O2的清除剂, DMTU、ASA和NaHCO3实验浓度参考文献[5, 7, 9]并由预试验筛选确定。为避免碱冲击幼苗, NaHCO3浓度每天按25 mmol· L-1递增至终浓度75 mmol· L-1后记为开始试验处理。叶面喷施于每天8:00和19:00进行, 喷施量以叶面滴液为限, 每盆喷施量约10 mL, 为降低表面张力, 喷施液中加2滴吐温-80。根部浇灌每天19:00进行, 浇灌量800 mL。处理7 d后取幼苗叶片用液氮速冻后-70 ℃保存, 用于测定相关生理指标。每5盆为1个重复, 重复3次, 随机排列。
幼苗鲜重测定采用称重法[5]; 超氧阴离子(
采用Excel 2003处理数据和绘图, SPSS 20.0方差分析和Duncan法多重比较(P< 0.05)。将所检测的16项指标测定值离差标准化后进行SPSS 20.0主成分分析, 结果提取4个主成分, 方差贡献率分别为57.155%、21.529%、8.393%和7.182%, 累积贡献率达94.260%, 计算得出4个主成分的得分值后按王玲等[16]的方法计算隶属函数值
U(Xj), 以方差贡献率占累积贡献率的比重作为权重Wj, 分别为0.606、0.228、0.089和0.076, 按公式D=∑ [U(Xj)× Wj]计算出各处理的综合评价值(D)。
燕麦幼苗三叶一心期进行75 mmol· L-1 NaHCO3胁迫, 取幼苗倒2叶检测H2O2含量显示, 当胁迫9 h时H2O2出现爆发式骤增, 随即急剧降低后转向逐渐升高(图1A)。H2O2“ 猝发” 说明其可能作为一种信号参与碱胁迫响应, 随胁迫时间延长H2O2又快速积累可能是碱胁迫伤害的表现。燕麦幼苗在75 mmol· L-1 NaHCO3胁迫的同时喷施0.001~5 mmol· L-1 H2O2 7 d后发现, 0.001~1 mmol· L-1 H2O2能够提高燕麦幼苗鲜重, 其中0.01 mmol· L-1 H2O2效果最佳(图1B)。因此, 以0.01 mmol· L-1 H2O2作为燕麦响应碱胁迫的实验浓度。
 | 图1 75 mmol· L-1 NaHCO3胁迫下燕麦幼苗叶片内源H2O2含量变化(A)及喷施不同浓度H2O2对幼苗鲜重的影响(B) CK:Control; T0:75 mmol· L-1 NaHCO3; T1:75 mmol· L-1 NaHCO3+0.001 mmol· L-1 H2O2; T2:75 mmol· L-1 NaHCO3+0.01 mmol· L-1 H2O2; T3:75 mmol· L-1 NaHCO3+0.1 mmol· L-1 H2O2; T4:75 mmol· L-1 NaHCO3+1 mmol· L-1 H2O2; T5:75 mmol· L-1 NaHCO3+5 mmol· L-1 H2O2.不同字母表示处理间差异显著(P< 0.05), 下同。Fig.1 Change of endogenous H2O2 contents in leaves (A) and effects of spraying 0.001-5 mmol· L-1 H2O2 for 7 days on fresh eight of oat seedlings under 75 mmol· L-1 NaHCO3 stress (B) Different letters indicate significant difference among the treatments (P< 0.05). The same below. |
从表1可知, NaHCO3胁迫(处理T1)引起了燕麦叶片
| 表1 喷施0.01 mmol· L-1 H2O2对75 mmol· L-1 NaHCO3胁迫下燕麦幼苗叶片 |
图2表明, 与CK相比, NaHCO3胁迫(处理T1)下燕麦幼苗叶片SOD、APX活性和ASA、GSH含量显著降低, CAT和POD活性明显升高; 喷施H2O2(处理T2)提高了NaHCO3胁迫下SOD、APX活性和ASA、GSH含量, 降低了CAT、POD活性; 添加DMTU处理(T4)与处理T2相比SOD、CAT、POD、APX活性和ASA、GSH含量均显著降低; 施用ASA处理(T5)与处理T2相比SOD、CAT、POD、APX活性和GSH含量显著降低, 而ASA含量却明显升高; 单独喷施H2O2(T3)与CK相比, SOD、APX活性和GSH含量降低, CAT、POD活性提高, ASA含量变化不大。
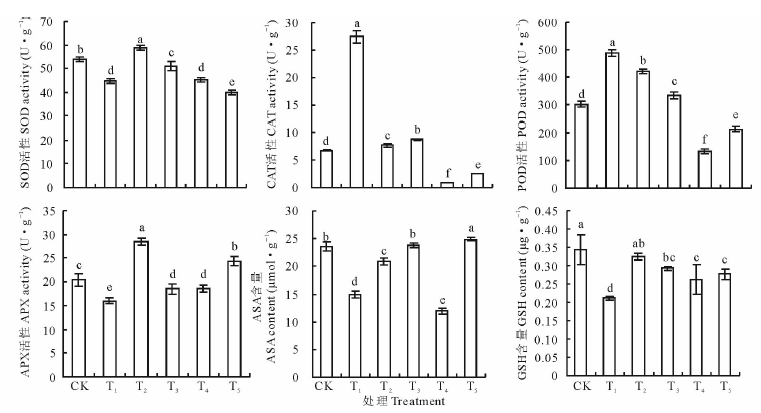 | 图2 75 mmol· L-1 NaHCO3胁迫下外源H2O2对燕麦幼苗叶片抗氧化系统的影响Fig.2 Effect of spraying 0.01 mmol· L-1 H2O2 on antioxidant system in leaves of at seedlings under 75 mmol· L-1 NaHCO3 stress CK:Control, 喷水spraying H2O+根部浇水root application H2O; T1:喷水spraying H2O+根施root application 75 mmol· L-1 NaHCO3; T2:喷spraying 0.01 mmol· L-1 H2O2+根施root application 75 mmol· L-1 NaHCO3; T3:喷spraying 0.01 mmol· L-1 H2O2+根部浇水root application H2O; T4:喷spraying 0.01 mmol· L-1 H2O2+根施root application 75 mmol· L-1 NaHCO3+根施root application 5 mmol· L-1 DMTU; T5:喷spraying 0.01 mmol· L-1 H2O2+根施root application 75 mmol· L-1 NaHCO3+根施root application 2 mmol· L-1 ASA. 下同The same below.14 |
由表2可知, 与CK相比, 碱胁迫(处理T1)提高了燕麦幼苗叶片可溶性糖、可溶性蛋白质、游离氨基酸和脯氨酸含量, 有机酸含量明显下降; 喷施H2O2(处理T2)降低了碱胁迫下可溶性糖、游离氨基酸和脯氨酸含量, 提高了有机酸含量, 可溶性蛋白质含量无显著变化; 施用H2O2淬灭剂DMTU处理(T4)与处理T2相比可溶性糖、可溶性蛋白质、游离氨基酸、脯氨酸含量均显著提高, 但有机酸含量明显下降; 施用H2O2清除剂ASA处理(T5)与处理T2相比可溶性糖、可溶性蛋白质、游离氨基酸、脯氨酸含量也是显著提高, 而有机酸含量明显下降; 单独喷施H2O2处理(T3)与CK相比, 可溶性糖、可溶性蛋白质、游离氨基酸含量都显著提高, 脯氨酸含量明显下降, 而有机酸含量变化不大。
| 表2 75 mmol· L-1 NaHCO3胁迫下喷施0.01 mmol· L-1 H2O2对燕麦幼苗叶片渗透调节物质含量的影响 Table 2 Effect of spraying 0.01 mmol· L-1 H2O2 on osmotica content in oat eedling leaves under 75 mmol· L-1 NaHCO3 stress |
由图3A可知, 与CK相比, NaHCO3胁迫(处理T1)显著降低了燕麦植株鲜重, 降幅达36.4%。喷施H2O2(处理T2)显著缓解了NaHCO3胁迫燕麦幼苗鲜重的降幅, 增添DMTU(处理T4)和ASA(处理T5)完全逆转了H2O2对NaHCO3胁迫下燕麦幼苗鲜重下降的缓解作用; 单独喷施H2O2(处理T3)与CK相比显著提高了燕麦幼苗鲜重, 增幅为15.5%。
对检测的
 | 图3 75 mmol· L-1 NaHCO3胁迫下喷施0.01 mmol· L-1 H2O2对燕麦幼苗鲜重的影响及其耐碱性综合评价Fig.3 Effects of spraying 0.01 mmol· L-1 H2O2 on fresh weight of oat seedlings under 75 mmol· L-1 NaHCO3 stress and comprehensive evaluation to alkali resistance of oat seedlings |
Fedina等[17]研究发现, 外源H2O2预处理能显著降低盐胁迫下大麦(Hordeum vulgare)幼苗MDA和内源H2O2含量。Abdul等[8]指出, 用H2O2浸种能够通过缓解氧化伤害提高小麦幼苗的耐盐性[8]。本试验表明, 碱胁迫下燕麦幼苗发生H2O2“ 猝发” 现象(图1A), 喷施H2O2能够有效抑制碱胁迫燕麦叶片
积累渗透溶质是燕麦适应碱胁迫的重要特征[5]。可溶性糖和脯氨酸等是细胞主要的有机渗透溶质。本研究表明, 喷施H2O2降低了碱胁迫下燕麦叶片的可溶性糖、游离氨基酸和脯氨酸含量, 却提高了有机酸含量; H2O2淬灭剂或清除剂逆转了H2O2的这种调节作用(表2)。表明H2O2参与燕麦幼苗耐碱性适应过程渗透溶质积累的调控。这与外源H2O2预处理可降低NaCl胁迫下大麦叶片脯氨酸含量的结果一致[18], 但与张波等[19]的外源H2O2提高盐胁迫下小麦可溶性糖含量的结果不同。说明不同胁迫下植物种类不同积累渗透溶质的机制存在差异。喷施H2O2降低碱胁迫下燕麦可溶性糖含量可能是其作为信号参与调节气孔关闭[7]后影响光合作用所致, 游离氨基酸和脯氨酸含量下降可能与喷施H2O2后降低碱胁迫燕麦内源H2O2含量(表1)使蛋白质降解下降有关[20], 但H2O2促进碱胁迫下有机酸积累的机制有待进一步探讨。
生物量是燕麦耐碱性强弱的直接指标, 也是生理响应的综合体现[5]。植物生物量与参与光合作用光能吸收和传递的叶绿素含量密切相关[1]。本试验表明, 喷施H2O2有效抑制了碱胁迫下燕麦幼苗叶绿素含量(表1)和植株鲜重(图3A)的降幅, H2O2淬灭剂或清除剂完全逆转了H2O2的作用。这与Azevedo等[21]的外源H2O2能够缓解盐胁迫玉米(Zea mays)幼苗生长抑制的结论一致。说明外源H2O2能够增强燕麦幼苗的耐碱性。这可能一方面归因于H2O2对燕麦幼苗SOD、APX活性和ASA、GSH含量的提升作用(图2), 使碱胁迫产生的活性氧能够快速地被清除, 从而阻止了对细胞膜脂的过氧化伤害(表1); 另一方面, 可能与H2O2参与碱胁迫下燕麦幼苗渗透溶质积累的调节有关(表2), 尤其是对有机酸积累的促进作用可能对增强燕麦适应碱胁迫造成的高pH胁迫发挥了重要作用。为了综合评价喷施H2O2对燕麦幼苗耐碱性的影响, 采用主成分分析提取了4个独立的综合性指标替代所检测的16项指标, 方差累积贡献率达94.26%, 对4个主成分得分值采用隶属函数分析综合评价耐碱性结果显示, 喷施 H2O2显著提高了碱胁迫下燕麦幼苗的综合评价值, H2O2的作用可被H2O2淬灭剂或清除剂部分或完全逆转(图3B), 这与H2O2对生物量的影响基本一致(图3A), 充分证明外源H2O2能够提高燕麦幼苗对碱胁迫的适应能力。
喷施0.01 mmol· L-1 H2O2能够显著降低75 mmol· L-1 NaHCO3胁迫下燕麦叶片中
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|